The brute force of “Why is there something rather than nothing?”
[Series Outline/Table of Contents]
Question 7: Chemistry: active bio-information and “struggle to survive”
====================================================================
Source notes—references below by
keyword refer to these multiple-quoted sources:
·
[Adami]
= The Evolution of Biological Information: How Evolution Creates Complexity,
from Viruses to Brains. // Adami, Christoph // PrincetonUP:2024. Kindle.
·
[Arias,
Master] = The Master Builder: How the New Science of the Cell Is
Rewriting the Story of Life . Alfonso Martinez Arias // Basic Books:2023.
Kindle.
·
[Davies,
Demon] = The Demon in the Machine --How hidden webs of information
are solving the mystery of life. // Davies, Paul // Penguin:2019.
·
[Denton,
Miracle] = The Miracle of the Cell. // Denton, Michael //
DiscoveryInstitute:2020.
·
[Koonin]
= The Logic of Chance: The Nature and Origin of Biological Evolution. //
Koonin, Eugene // FTpress:2012.
·
[Living
Systems] = Understanding Living Systems. // Noble, Raymond and
Noble, Denis. // Cambridge University Press. Kindle Edition.
·
[Lynch,
Origins] = The Origins of Genome Architecture. // Lynch, Michael
// OxfordUP:2018.
·
[Prebiotic]
= A Primer in
Prebiotic Chemistry.
// Albert C. Fahrenback and Henderson J Cleaves II // OUP:2024.
·
[Purpose] = Evolution "On Purpose":
Teleonomy in Living Systems (Vienna
Series in Theoretical Biology). // Corning, Peter A.; Kauffman, Stuart A.;
Noble, Denis; Shapiro, James A.; Vane-Wright, Richard I. // MIT Press:2023.
Kindle Edition
·
[Shapiro]
= Evolution: A View from the 21st Century. Fortified. //Shapiro, James
A. // Cognition Press:2022. Kindle
Edition.
·
[Transformations]
= Transformations of Lamarckism: From Subtle Fluids to Molecular Biology (Vienna
Series in Theoretical Biology), // Snait B. Gissis, Eva Jablonka, and Anna
Zeligowski . // MIT:2011. Kindle Edition.
·
[Transitions]
= The Major Transitions in Evolution Revisited (Vienna Series in
Theoretical Biology). // Brett Calcott and Kim Sterelny, // MIT:2011
·
[Walker]
= Life as No One Knows It: The Physics of Life's Emergence. // Walker,
Sara Imari. // Penguin Publishing Group:2024. Kindle Edition.
[Also—for
practical and navigation reasons--when quoting source material, I have
generally omitted THEIR use of sources, as they would include that in
(parentheses). But—for the interested reader who wants to drill down on the
sources’ sources, I have indicated WHERE they had referenced sources by LEAVING
the parentheses, but REMOVING the contents. So, for example, (Goodman and
Dorsey, 1948) became simply (---) in the quote. The interested reader could
find the source of MY quote and then inspect it for THOSE (omitted)
references.]
[AI
usage disclosure: All of the text in this article was written by me, or by the
authors of the individual quotes. I use AI (Claude) to summarize one chart, and
identified it as being from Claude. I DID use AI heavily in producing the
pretty charts (as a favor to the readers – smile). Although all the data used
in the charts was reviewed and checked – and several iterations were required
to verify them.]
…………………………………………………….. ………………………………………..
Question 7: Chemistry: active bio-information and “struggle to survive”
“Why -- rather than nothing–
-- is there
something – so ‘law-like’ , such that physics is possible,
-- of a scale that
is incomprehensible in itself,
-- containing
levels of complexity and patterns that stagger the mind,
-- that looks structured
to allow ‘life’ as we know it, and
-- that contains
uber-complex chemical structures / assemblies that contain chemical sub-units that are so finely-tuned with OTHER chemical
structures that they REPEATEDLY ‘act on’
one another, and with one another, in a coordinated fashion to maintain
the whole, operate on a shared ‘clock’ in doing this, cooperate to respond to internal threats, reproduce themselves, respond
to external conditions (e.g. threats,
energy sources) as a unit, and occasionally MODIFY THEMSELVES to fulfill
these requirements—when all other assemblies of chemical structures
DO NOT, and when we have no real
explanation of how they got here nor an understanding of why they are STILL
HERE!
Up to this point, “all” we have
been confronted with is a universe (subset of SOMETHING) that has the building
block elements necessary for life—but which are not alive themselves, and which
actually do NOTHING without some ‘mixing and stirring up’.
And, because of ‘timing’ of cosmic
factors, we have these building blocks in a narrow window of the universe in
which ‘life’ COULD appear (and be sustained for a while).
But once our species ‘woke up, we
noticed the amazing biosphere — we were surrounded by ‘living things’. And
during our history, we discovered that there were single-celled living things
that had been around much, much longer than we had, and which—without a ‘brain’
or ‘nervous system’—did all of the actions mentioned above—as ‘living cells’.
“The cell is a
microscopic reaction vessel—a tiny ‘flask’. Despite its small size (and
possibly because of it), a cell can carry out
thousands of different reactions simultaneously, in a coordinated
and highly regulated way, connected through positive and negative feedback
mechanisms so that the cell is able to manufacture what it requires
responsively. These feedback mechanisms allow the cell to behave as though it ‘knows what it’s doing’,
responding to the environment to optimize its chances of success and survival,
ultimately amplifying its own reproduction while relying on just a few basic
inputs from the environment.” [Prebiotic pp.3-4]
We note here how ‘odd’ this ‘life’
is – maybe even worse in implications than the previous series of
‘oddness’.
I use the ‘maybe worse’ phrase
deliberately. If ‘something’ itself ‘(the well-ordered universe we see) should
not be/exist’ (nothing ‘should be’ the default, remember), then “life”
as we see it now is does not even FIT here…
Life as we experience it now SEEMS
to be built from all the physics and chemistry stuff, but SEEMS to be radically
DISCONTINUOUS with that…
If you look at a jar of wiggling bacteria
and then look at another jar containing the exact same amounts and sequences of
chemicals that are in the bacteria—just ‘disassembled’ and mixed up—you would
NOT assume they were related in any way.
Life just does not seem to ‘fit’ in
a well-ordered and (reasonably) predictable physical universe.
·
Actions
in physics ‘move’—but always on their way to eventual thermodynamic
flat-lining.
·
Actions
in chemistry ‘move’—but always on their way to equilibrium’s
‘stillness’. [auto-catalysis notwithstanding]
·
Actions
in life ‘move’ but NEVER reach equilibrium and CONSTANTLY ‘outrun’ the 2nd
law of thermodynamics! Life runs ‘above’ those powerful processes.
But when a cell DIES, its corpse
then takes its place in those natural, physical
pathways, decomposing and giving up all its stored energy to its
surroundings.
“These metabolic
reactions almost never reach equilibrium; in fact, if equilibrium in a
cell were ever reached, then one might say the cell was dead.” [Prebiotic,
p4.]
“One hint as to
where a missing element may lie comes about from a simple observation. A living
thing is not just a particular physico-chemical assembly, an aggregate of
various molecular components synchronously functioning together like the cogs
and wheels within a mechanical watch. A living system is better understood as
an energized physico-chemical assembly (---), one which both stores and depends
on a continuous supply of material and energy for its maintenance. If the
material and energy supplies are cut off, the living system ceases to exist—it
dies. Only then does that chemical aggregate
behave as chemical aggregates normally do. That explains why life is
so difficult to create. Simply, we still do not know how to generate dynamic
energized systems of that kind.” [Purpose, pp. 259-260]
The seeming disconnect between
living things and the non-living things that are USED by living things is so
stark -- to me -- that it seems almost like another ‘something from nothing’
problem. [Not as severe as the main version perhaps, but still without any
PLAUSIBLE origination process (efforts and spot successes in the OOL research
area notwithstanding)]. It just ‘appears’ in geologic history, without any
record of ancestry and without (so far) any demonstration in labs of PLAUSIBLE
ancestry (brilliant work, IMO, but widely recognized as not-even-close-yet…).
…………………………
But to get back to base…
My descriptive statement above
contains these phrases:
1.
That
contains uber-complex chemical structures / assemblies, containing chemical
sub-units that are so finely-tuned with OTHER chemical structures that they
a.
REPEATEDLY
‘act on’ one another, and with one another, in a coordinated fashion to
maintain/grow the whole,
b.
Operate
on a shared ‘clock’ in doing this,
c.
Cooperate
to respond to internal threats (e.g.
DNA and cellular repair mechanisms)
d.
Reproduce
(when it senses that conditions are right for that)
e.
Respond to external conditions (e.g. threats, energy
sources) as a unit, and
f.
Occasionally MODIFY THEMSELVES as needed to fulfill
the requirements above…
2.
When
all other assemblies of chemical structures DO NOT…
3.
And
when we have no real explanation of how they got here…
4.
Nor
how they are STILL here!
I am talking about the units we
call ‘living CELLS’ , ranging from simple single cells (e.g. archaea,
bacteria), generally (but not exclusively—see next question in the series)
living independently of other cells, as well as cells as part of a
multi-cellular unit (e.g. eukaryotes).
Nobody on the planet really knows
what ‘life’ is, but most of the working definitions by researchers include
these basic requirements:
1.
It ‘eats’ and grows: Metabolism (coordinated
processes to sustain activity)
2.
It produces offspring: Information-based
Reproduction/replication (e.g. DNA)
3.
It watches what is happening, and makes adjustments to the above as
needed: Sensory and signaling mechanisms to detect/respond to threats
(both internal and external threats).
My ‘characteristics’ list would
fall into those categories like this:
·
Metabolism
o
“Repeatedly
acting upon one another, and with one another, in a coordinated fashion
to maintain the whole (would include internal growth and activity, digestion
and waste processing; renewal of internal components—e.g. making proteins and
managing energy resources)”
o
“Operating
on a shared ‘clock’”
·
Information-based Reproduction/replication
o
“Reproduce
themselves (when it senses that conditions are right for that)”
·
Sensory / signaling mechanisms to detect/respond
o
“Cooperate
to respond to internal threats”
o
“Respond
to external conditions (e.g. threats, energy sources) as a unit”
o
“Occasionally
MODIFY THEMSELVES to fulfill these requirements”
The ACTIONS I describe below fall
into one or more of those categories.
Okay, let me go through these
elements, and give a piece of data or two, and perhaps a representative example
or two…
ONE: uber-complex
chemical structures / assemblies within the cell, as seen in SIZE, CONTENTS,
and ACTIONS…
SIZE: The first thing to
note here is how ‘odd’ the SIZE gap
is between inorganic molecules and the smallest ‘life’ units
(e.g. phages, virus, bacteria).
The largest
naturally occurring INORGANIC molecules on record currently are packratite,
vanarsite, ophirite, and morrisonite, ranging between 5.75kDa
and 5.8kDa.
·
The
smallest semi-living unit (the MS2 bacteriophage, a virus)
has a single-stranded RNA molecule consisting of 3,569 nucleotides. At 330
Daltons per nucleotide, that sums up to 1.18MDa.
·
That
single molecule is 202 times larger than packratite.
·
The
total weight of the virus (since it contains 180+ proteins too)
is 3.91MDa, which is 660 times larger than packratite.
This size gap gets
much wider when we move to things that are classified as ‘fully alive’
The smallest cells
that are considered to be alive, fall into 3 major categories:
1.
Obligate
endosymbiont-- they are alive but
are dependent on other organisms for some of their cellular
functions.
2.
Parasitic symbiont– can reproduce and act
independently, but mostly thrive inside hosts
3.
Free-living (able to live and reproduce independently
of other cells/organisms)
(Types 2 and 3 we
will discuss in the next ‘shock question’ – the social ontology of life as we
know it.)
The smallest
organisms found so far in these categories are these:
·
Obligate
endosymbiont: Nasuia deltocephalinicol
·
Obligate
endosymbiont: Nanoarchaeum equitans
·
Parasitic
symbiont: Mycoplasma genitalium
·
Free-living:
Pelagibacter ubique
When you compare
the size of the largest inorganic molecule to the sizes of just the one chromosome
molecule in each of these cells, the gap is a chasm:
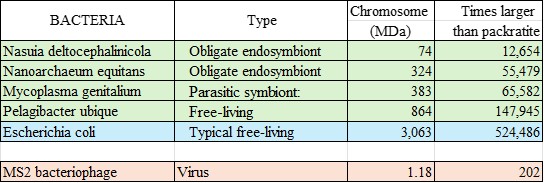
The smallest
living thing has a molecule over 12,000 times larger than
packratite.
And our familiar ‘lab
partner’ e.Coli’s genome is over half-a-million
times larger.
This size gap is
almost shocking—why are there NO objects in between
these sizes?
Here is a chart of
unit masses (not just molecules)—on a log scale!

Authorities
recognize that the amount of COMPLEXITY of even the simplest of cells, requires
SO MANY MOVING PARTS that it just cannot be reduced in size to that extent:
“Though the process by which life emerged continues to
mystify, there is one facet of the process that is beyond
doubt: the extraordinary degree of complexification that took place during that
extended physico-chemical transformation. The conversion of some
prebiotic chemical system, though of unknown identity, into simplest life would
have involved an increase in size/mass of
some nine orders of magnitude. [Molecular systems are typically
of mass of ca. 10(-21) g while that of a bacterium is ca. 10(–12)
g]. That’s a staggering change in both size and structural complexity—the
equivalent of something the size of a coin growing into something the
size of our planet. In fact, it seems reasonable to conclude that
life’s unique nonmaterial characteristics must have emerged as a direct
consequence of that extraordinary degree of complexification (Pross, 2013). So
how can that process of complexification be explained? [Purpose, p. 259]
Although there are
no ‘transitional forms’ in the record, the presumed path from non-life
to life would have had to start with something AT LEAST 100 times larger
than packratite.
Koonin [Koonin,
p. 438] calculates values for a simplified ‘toy’ model, that deliberately
over-inflates the rate of RNA replication and comes up with this statement:
“The requirements for the emergence of a primitive,
coupled replication-translation system, which is considered a candidate
for the breakthrough stage in this paper, are much greater. At a minimum, spontaneous
formation of the following is required:
• Two rRNAs, with a total size of at least 1,000
nucleotides.
• Approximately 10 primitive adaptors of about 30
nucleotides each, for a total of approximately 300 nucleotides.
• At least one RNA
encoding a replicase, about 500 nucleotides (low bound) required.
This totals out at
1,800 nucleotides, and at 330Da per, gives 594kDa as the smallest
possible proto-life unit.
That is 100
times larger than packratite—and that is only the replicator
‘half-way’ point to what Koonin calls ‘bona fide biological
entities’…
CONTENTS: Even
the simplest of cells
are filled with molecules in complex arrangements, of a wide variety of types,
and with complex interactions—from Day One.
The oldest life
forms that leave a fossil trace of its life in the geological record is
reported as being around 3.2 BYA. And evidence of simpler life forms (stromatolites,
but not fossils) is reported around 3.5 BYA.
“According to one
view, a diverse assemblage of microbes, including cyanobacteria-like cells,
had colonized the seas by -3.5 BYA (---). Some aspects of this interpretation
of the fossil record have been questioned (---), but other signs of biological
activity have been found in rocks from 3.4-3.8 BYA (---), and unambiguous
fossils of filamentous organisms deposited around hydrothermal vents
have been dated to 3.2 BYA (--). Thus, cellular life (as we know it)
appears to have emerged from inorganic materials within a window of just a few
hundred million years. [Lynch,
Origins,
p.2]
“The lack of
distinctive morphological features renders the detailed taxonomic composition
of the early fossil record quite uncertain, but the first 0.5-1.0 billion
years of life appear to have been dominated by prokaryotes (simple
cells lacking membrane-enclosed organelles, often referred to as bacteria), if
not entirely restricted to them. … The first evidence of eukaryotes
(cells with membrane-enclosed organelles, including a nucleus) appears in the
form of putative diagnostic biomarkers of membrane components deposited in
shale from -2.7 BYA (---). The first presumptive algal fossils date to -2.1 BYA
(---), and many other fossils of unicellular eukaryotes with well-developed
cytoskeletons date to 1.7-1.5 BYA (---). But despite this gradual addition of
eukaryotic diversity, the biosphere continued to be dominated by unicellular
and oligocellular (a few cell types) species for at least another billion
years.” [Lynch, Origins, p.2]
And
“Stromalites are
macroscopic sedimentary structures formed by microbial communities. As these
communities grow, they deposit thin layers of (generally) calcareous (calcium
based) minerals, or trap and bind calcareous sand grains, forming a stratified
structure… Fossilized stromatolites appear through the fossil record, and
indeed are the only fossil for almost 3 billion years of Earth history,
but the oldest ones that have general scientific consensus date back to nearly 3.5
billion years ago, and are also found in Western Australia. … Taking the appearance of liquid
water as a necessary condition which can be constrained at 4.2 billion years
ago, we can estimate that life emerged on Earth somewhere between 4.2 and at
least 3.5 billion years ago—a 700 million-year window.” [Prebiotic, pp. 34-35]
These earliest life forms were already very complex and sophisticated:
“Given the
presence of methanogenesis as early as 3.0BYA and photosynthesis as early as
2.8 BYA, we can conclude that the microbial
world was quite sophisticated biochemically by this time, perhaps
harboring the full repertoire of metabolic /molecular processes from which all
subsequent cellular lineages were built.” [Lynch, Origins, p.2]
We can check this
remark about ‘quite sophisticated’ by noting how the fossils were
interpreted:
“Here we
report the discovery of a population of large (up to about 300 micrometers in diameter) carbonaceous
spheroidal microstructures in Mesoarchaean shales and siltstones of
the Moodies Group, South Africa, the Earth's oldest siliciclastic alluvial to
tidal-estuarine deposits. These microstructures are interpreted as organic-walled microfossils on the basis of
petrographic and geochemical evidence for their endogenicity and syngeneity,
their carbonaceous composition, cellular morphology and ultrastructure,
occurrence in populations, taphonomic features of soft wall deformation, and
the geological context plausible for life, as well as a lack of abiotic
explanation falsifying a biological origin. These are the oldest and largest
Archaean organic-walled spheroidal microfossils reported so far. Our
observations suggest that relatively large
microorganisms cohabited with earlier reported benthic microbial
mats in the photic zone of marginal marine siliciclastic environments 3.2
billion years ago. “
[Javaux EJ,
Marshall CP, Bekker A. “Organic-walled microfossils in 3.2-billion-year-old
shallow-marine siliciclastic deposits”. Nature. 2010 Feb
18;463(7283):934-8.]
Notes:
1.
A
300 micrometer cell is 150-300 times larger than e.Coli…
2.
These
were interpreted early as being versions of chemotrophic beggiatoacean gliding
bacteria (can get up to 200 micrometers) and with photoautotrophic
oscillatoriacean cyanobacteria (up to 10 micrometers).
3.
Schopf
disputed the photo-synthetic interpretation and compared them to non-photosynthetic cyanobacterium-like cells.
4.
If
they are in the cyanobacteria family (photo or not) they were already
VERY sophisticated: “Cyanobacteria are challenged by environmental
stresses and internally generated reactive oxygen species that cause DNA
damage. Cyanobacteria possess numerous E.
coli-like DNA repair genes.[--] Several DNA repair genes are highly
conserved in cyanobacteria, even in small genomes, suggesting that core DNA repair processes such as recombinational
repair, nucleotide excision repair and methyl-directed DNA mismatch repair are
common among cyanobacteria.”
5.
They have a wide range of genome sizes,
ranging from 1,199 to 8,383 protein-coding genes (average of 3,580).
[Zamani Dahaj,
Seyed Alireza. (2015). “A phylogenetic model to predict the patterns of
presence and absence of genes in bacterial genomes and estimate the frequency
of horizontal gene transfer.”]
6.
By comparison, e.Coli averages around
3,850 – so this is a large, complex cell – with no precursors in the fossil record
so far….
And the current
versions of these ‘simplest’ life forms are still ‘stupendously complex’:
“Whatever the minimal
complexity for life may be, there is no doubt that even the simplest known life
form is already stupendously complex. Indeed, life’s complexity is
so daunting that it is tempting to give up trying to understand it in
physical terms. A physicist may be able to give an accurate account of a
hydrogen atom, or even a water molecule, but what hope is there for describing
a bacterium in the same terms?” [Davies, Demon, pp. 82-83]
“I’m fascinated
by what makes organisms tick, what enables living matter to do such astounding
things – things beyond the reach of non-living matter. Where does the
difference come from? Even a humble bacterium
accomplishes things so amazing, so dazzling, that no human engineer can match
it. Life looks like magic, its secrets cloaked by a shroud of
impenetrable complexity. Huge advances in biology over the past decades have
served only to deepen the mystery. What gives living things that enigmatic
oomph that sets them apart from other physical systems as remarkable and
special? And where did all this specialness come from in the first place?”
[Davies, Demon, pp. 1-2]
“The simplest bacteria or archaea are also incredibly complex structures, typically containing thousands of genes and complex
metabolisms with intricate control mechanisms. It remains challenging to
understand how something as seemingly simple as a single-celled organism arose.
It is thought that the first cells, sometimes referred to as protocells, must
have been much simpler than even the simplest modern prokaryote.” [Prebiotic,
p.2]
Let’s get a visual
on this ‘gulf’ between non-life and life, starting with ‘how much stuff’ is
inside even the smallest of living/semi-living cells:
Here’s a chart of
the number of ATOMS in the largest
INORGANIC molecule (Packratite), the smallest virus, the smallest living
cell (obligate), and the smallest free-living cell:
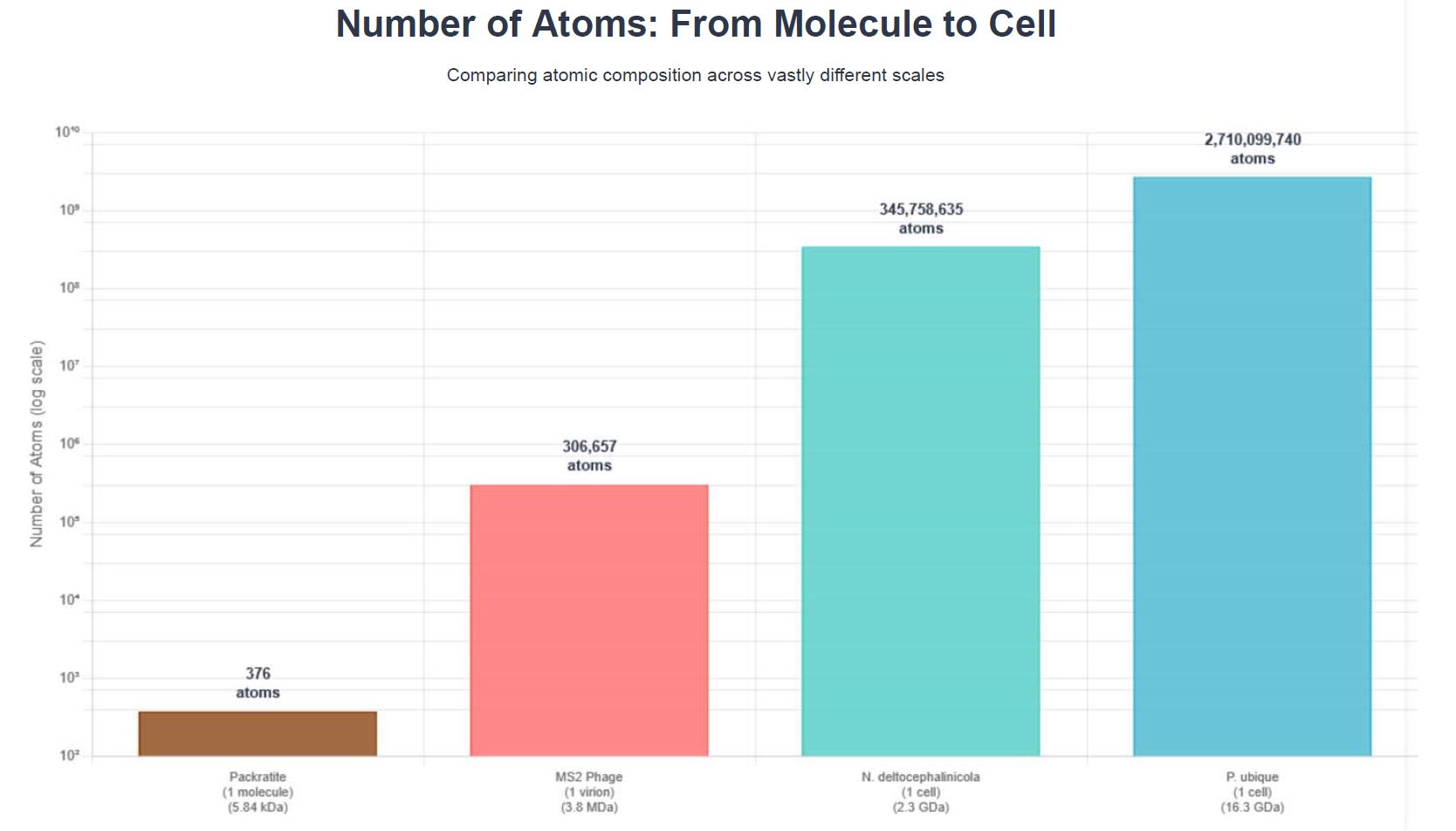
This chart is on
the log scale which means each ‘layer’ is an order of magnitude higher than the
previous one – e.g. 10 times greater.
Notice :
·
That
our inorganic molecule has 376 atoms, and the smallest virus has a thousand
times more atoms in it.
·
That
the smallest cell considered alive, has 1 million TIMES more atoms
inside it.
·
That
the smallest free-living cell has over 7 million TIMES more atoms inside
it.
For these tiny
cells to have so many elements inside them, gives a good sense that they are
certainly ‘complex’ in their makeup!
ATOMS inside these cells
are obviously combined into MOLECULES.
And their story is just as indicative of complexity as the other.
Here is a chart of
those smallest living things (plus E.
coli as reference). The bars represent the relative sizes of the cells, with the
organism’s name/type, and with the weight of that cell on the
right [“fg” stands for “femtogram” = 10(-15)
grams. A dry grain of rice weighs about 20 million femtograms.]
The number inside
the bars represents the average number of MOLECULES in that species’ cell.
So, the smallest living thing on the planet (that we know of) has
3.4 MILLION molecules in it.
We can even be
amazed at the SCALE of these components/cells, when compared to cosmic
phenomena. I selected only certain items for comparison—more could be done,
with similar results:
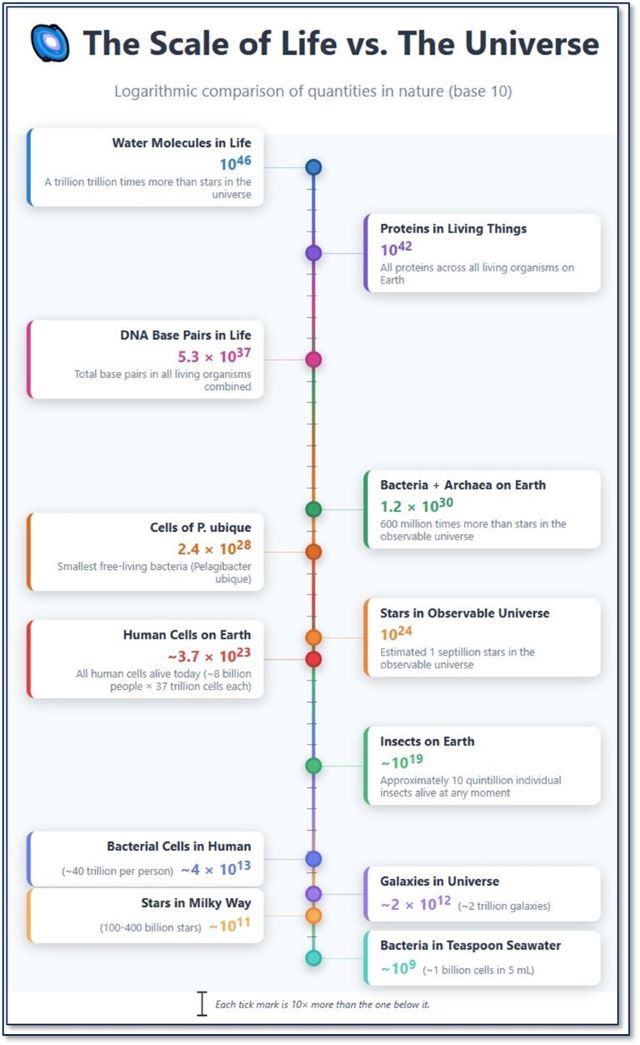
The summary of
this data offered by Claude-AI, highlights the scale differences:
“This logarithmic
chart reveals the staggering abundance of life's molecular components
compared to astronomical objects. The scale above spans from 109
to 1046 - that's 37 orders of magnitude!
“Key insights:
• All human cells
on Earth combined (~3.7 x 1023) are roughly comparable to the number
of stars in the observable universe (1024)
• There are more
insects on Earth (~1019) than there are galaxies in the universe (1012)
by a factor of 10 million
• A single human
body contains ~40 trillion bacterial cells (1013) - about 100 times
more bacteria than there are stars in the Milky Way
• A single
teaspoon of seawater contains about a billion bacteria.
• Water molecules
in living things outnumber stars in the universe by 1022 (10
thousand billion billion times)
• The entire
observable universe contains roughly 2 trillion galaxies, each with billions to
trillions of stars
“While the
universe is vast in terms of space, life is extraordinarily rich in complexity
and quantity at the molecular scale.
“This chart
reveals that even though individual organisms are tiny, the collective
molecular abundance of life on Earth rivals or exceeds the quantities we see at
cosmic scales. A single human is a walking ecosystem containing more bacterial
cells than stars in our galaxy!”
You should be able
to get an intuitive sense of how COMPLEX (and maybe ‘cluttered’?) these simple
life-forms are.
ACTIONS: It
is in this area that the uniqueness of the cell stands out so sharply.
These molecules
are not just ‘floating around’ inside some kind of pool inside the cell walls.
They are in constant motion and constant activity, being driven by the
‘decisions’ by the cell. They are agents in an endless cycle of metabolic
reactions that interface with other metabolic reactions, all monitored and
regulated by the cell itself.
We will get to the
interactions between them, in the step about ‘repeatedly ‘act on’ one another (i.e.,
metabolism), but let me give some data on HOW BUSY these little components are:
“Within cells,
there are enzymes that perform millions of chemical reactions each
second. Protein ‘motors’ make energy-storing molecules by spinning around thousands
of times a minute. Sugar molecules fly by at 250 miles per hour,
nearly double the speed of a Cessna 172 airplane at cruising speed. Every
protein in the cell is hit by 1013 water molecules every second.”
[Niko McCarty, Fast Biology, Asimov Press]
“… ATP synthase, a
barrel-shaped protein, made from three subunits, that makes the energy storage
molecules of the cell. ATP synthase does this by combining ADP with a free floating
phosphate to form ATP. The barrel-shaped protein sits embedded in a membrane
and performs this chemical reaction by harnessing the flow of protons down a
concentration gradient. It spins around and around, each rotation yielding
three molecules of ATP. Making ATP is, for these reasons, both a chemical
and a mechanical process. … The scientists observed that ATP synthase spins
around exactly 134 times each second, or 8,040 times each minute.
That speed is significantly faster than the propeller on most piston airplanes,
and about half the RPM of a Boeing 737 jet engine.”
[Niko McCarty, Fast Biology, Asimov Press]
“We
make use of one of our tricks of the trade which states that in E. coli a
single molecule per cell (say our substrate) has an effective concentration of
about 1nM (i.e. 10-9 M). The rate of collisions is thus [math
removed], i.e. they will meet within a second on average. This allows us to
estimate that every substrate molecule collides with each and every
protein in the cell on average about once per second. As a concrete
example, think of a sugar molecule transported into the cell. Within a second it
will have an opportunity to bump into all the different protein molecules in
the cell.” [Cell Biology by the Numbers, p. 275, Milo/Phillips/Orme.]
Note from above that there are 3,000,000
proteins in a single E. Coli cell.
“Another tiny bio-machine that has attracted a lot of
attention is a sort of freight-delivery molecule called kinesin. It
carries vital cargoes by walking along the tiny fibres which crisscross cells.
It does this gingerly – one careful step at a time – to avoid being swept away
by the incessant bombardment from the
thermally agitated water molecules that saturate all living cells and
move twice as fast as a jetliner.
One foot stays anchored to the fibre and the other comes from behind and sets
down ahead; then the process is repeated with the other foot. The anchor points
are where the binding forces between the foot and the fibre are especially
propitious: those sites are 8 nanometers apart, so each step is 16 nanometers
in length. It’s unnerving to think that billions of these little kinesin
wonders are creeping around inside you all the time. [Davies, Demon,
p.57]
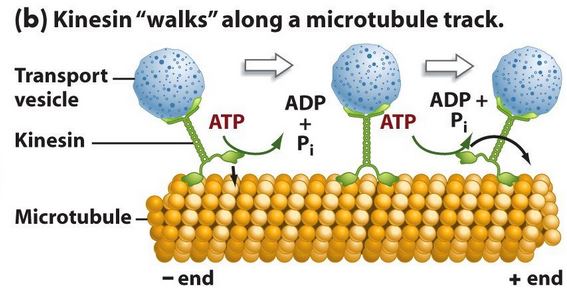
1A: REPEATEDLY ‘act on’ one another…
This is the amazing cellular
process called ‘metabolism’—in which these millions of chemical
reactions and processes run constantly, and produce outputs that are used as
inputs in OTHER chemical reactions. These overlapping and intertwining
processes work as a network, with control mechanisms and alternatives.
These ‘chains’ of chemical reactions are known as ‘pathways’, and there are a few that are considered ‘central’, because they produce both their OWN primary output, but also metabolites that FEED other pathways.
“While individual
organic reactions and the synthesis of specific compounds were undoubtedly
important for the origins of life, these must
have become organized into connected networks which helped convert
environmentally supplied compounds into the components of the network,
which stored and processed information about the network, catalysed the network
and helped maintain its components in close spatial proximity. … “The network of
reactions occurring within cells which affords the synthesis and breakdown of
the materials life needs (e.g. amino acid and nucleotide
building blocks), as well as the energy producing re-actions needed for general
cell maintenance also matter. Cells carry out an incredible number of
coordinated reactions in this manner. The biosynthetic pathways, which help
interconvert the material composition of the cell as well as provide energy for
cell function are collectively referred to as metabolism.” [Prebiotic,
p. 47]
“The chemical
reactions of metabolism are organized into specific pathways,
which are typically (but not always) catalyzed by protein enzymes. This
organization into pathways is crucial for allowing efficient capture and use of
the energy provided from and required for different transformations…While each
organism has its own particular set of metabolic reactions as a consequence of
its evolutionary ancestry, there are some core pathways which are common to nearly
all organisms, albeit with some modifications depending on the organism in
questions. In this chapter, we will briefly survey some of these central
prokaryotic metabolic pathways that provide precursor metabolites for other pathways.” [Prebiotic, p. 46]
The Primer on Prebiotic
Chemistry that I have been quoting from lists these as the central pathways
for Prokaryotes (archaea and bacteria);
·
Tricarbolic acid cycle (TCA, aka citric
acid or Krebs cycle). It produces the ‘energy packet’ ATP, and provides intermediates
for the lipid, amino acid, and nucleotide biosynthesis pathways.
·
Acetyl-CoA pathway. This produces Acetyl-CoA from
CO2.
·
Glycolysis. Breaks down sugar, giving ATP , and
supplies intermediates to other pathways.
·
Pentose phosphate pathway (PPP). This runs in
parallel with glycolysis and shares some of the same intermediates. It
is critical for providing the pentose phosphates need for DNA, RNA, and other
cofactors.
Other pathways (needed for synthesis
of other needed compounds):
·
Gluconeogenesis – opposite of glycolysis – rebuilds
sugars.
·
Amino Acid biosynthesis. Uses intermediates
from the TCA/Krebbs cycle to make amino acids.
·
Nucleotide biosynthesis. These are
multiple pathways, constructing RNA and DNA nucleotides.
·
Phospholipid biosynthesis. Creates needed
chains of lipid molecules.
Just to show you the complexity
of these, here are THUMBNAILS of the schematics for them.
Just these images will give you a
sense of how CONNECTED these things are – how they are constantly and
REPEATEDLY ‘acting upon’ one another.
And this is just for prokaryotes
as a group…
For the archaea sub-set of those,
here is a glimpse of just one of the pathways, the central metabolic pathway,
for one of the members:
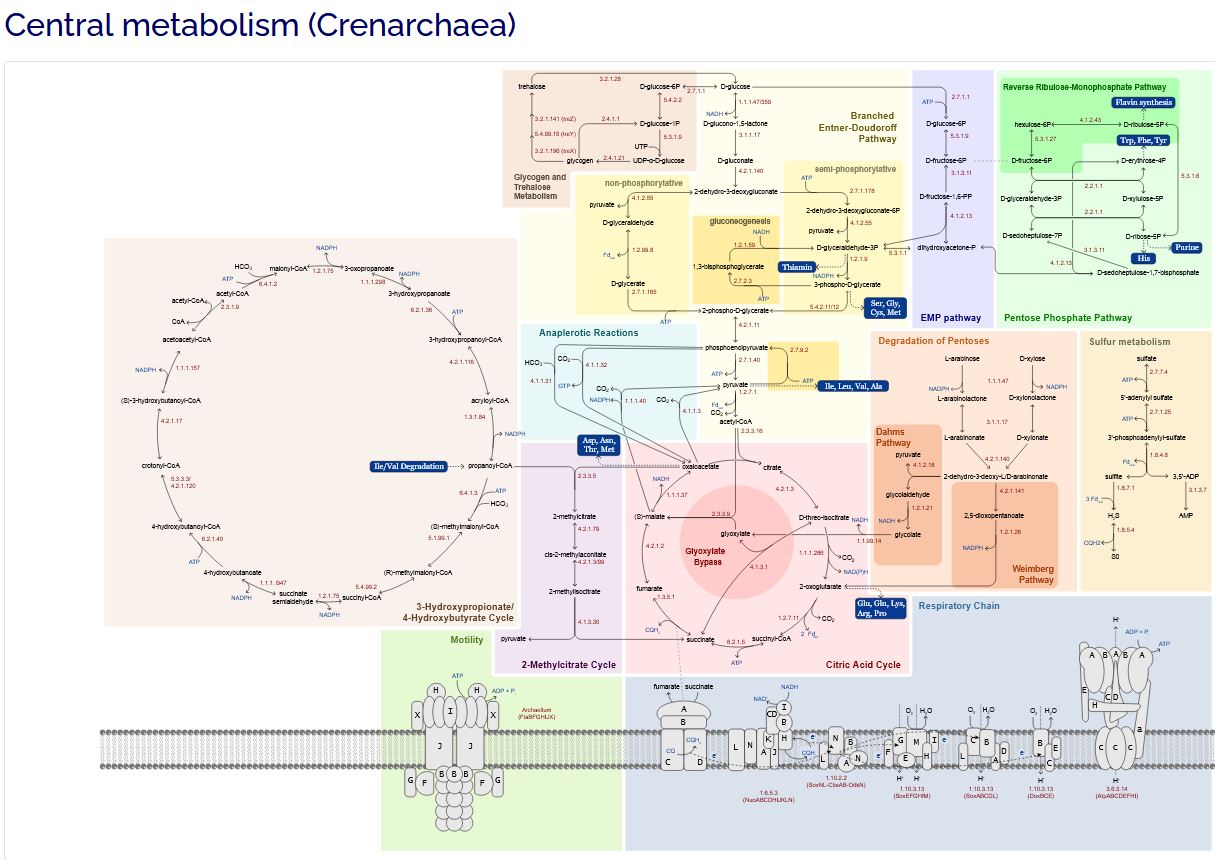
Each one of the nodes in those
maps/schematics represents more LEVELS OF DETAIL down (at least one).
[From the BRENDA
enzymes site, at the MetaboMAPS sub-site for pathways:
https://metabomaps.brenda-enzymes.org/ ]
[Also see the
Protein Data Bank, with visualizations of proteins and seqs:
There is an on-line ‘master’
schematic of all known (or possible) pathways, for most known life forms, posted
online, which can be populated with specifics by species, and interrogated by
pathway:
[ https://www.kegg.jp/kegg/pathway.html ]
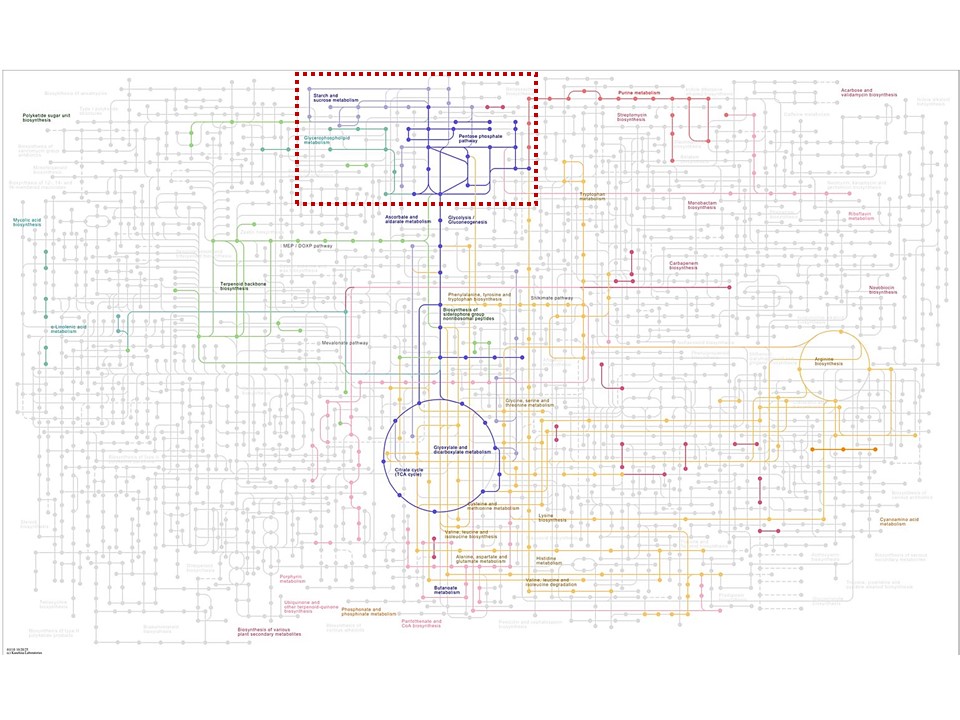
Here is the master template from
the site, with the lines representing pathways and the nodes representing
compounds:
Just for shock value to show the ‘organic depth’ of life– (lol) – here is the master schematic with some of e. Coli’s pathways selected:
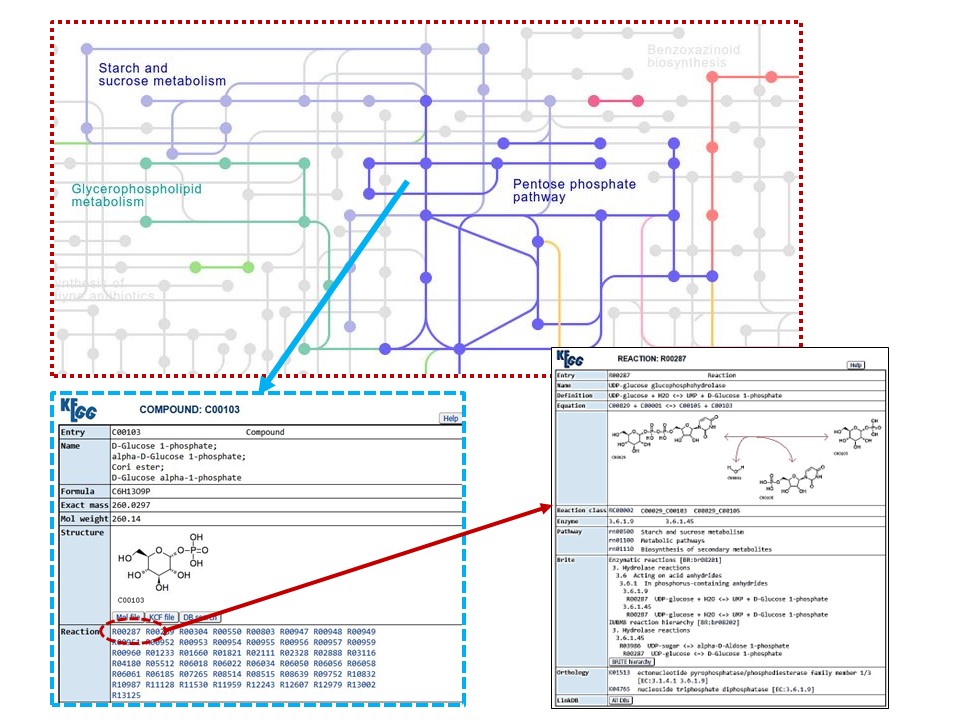
And here is the drill-down on the
red-dotted area. When clicking on a node in the chart, it opens up a window
describing the COMPOUND, and then there are links to all the REACTIONS it
participates in…the entire diagram is like this, revealing the deeper and
deeper levels of complexity (and sophistication) of the living cell:
This complexity is mind-boggling—especially
at that scale/size!—and seems so
out-of-place, compared to the levels of complexity ‘beneath it’ (i.e.
inorganic materials), and even—in some real sense—beyond
the complexity of the physical cosmos itself (as noted above).
More or less incomparable:
“Cells are
amazing. Even to a non-biologist, they convey the impression of being very
special objects with extraordinary capabilities. No one who has observed a
leucocyte (a white blood cell) purposefully—one might even say single-mindedly—chasing
after a bacterium in a blood smear would disagree. … What one witnesses
there seems to transcend all our intuitions: A tiny speck of matter,
invisible to the naked eye, so small that one hundred of them could be lined up
across the top of a pin, is seemingly endowed with intention and agency.
It's like watching a house cat chasing a mouse, or a cheetah chasing a gazelle
on the African savanna, or indeed a man chasing down a kudu in the Kalahari.
“It does not
lessen the amazement to conclude that this ability must arise somehow from the
atomic complexity that lies within this wondrous speck of matter. For the
complexity in which this behavior is instantiated is also far beyond ordinary
experience. A cell consists of trillions of atoms, representing the complexity
of a jumbo jet and more, packed into a space less than a millionth of the
volume of a typical grain of sand. But unlike any jumbo jet, unlike any nano-tech,
or indeed unlike even the most advanced human technology of any kind, this
wondrous entity can replicate itself.
Here is an "infinity machine" with seemingly magical powers…
“In terms of
compressed complexity, cells are without peer in the material world, actualized
or imagined…There is much more to discover about the cell, but even from our
current limited knowledge of its depths it is clear that this tiny unit of
compact, adaptive sophistication constitutes something like a third infinity.
Where the cosmos feels infinitely large and the atomic realm infinitely small,
the cell feels infinitely complex.”
[Denton, Miracle,
p16]
And the more that researchers
learn, the deeper the mysteries are,
and the more ‘poorly understood’ phrases
there are (which still amaze us in their sophistication):
“Only as the veil began to lift with the mid-century
molecular biological revolution did science begin to glimpse the
sophistication of these extraordinary pieces of matter. Subsequently, every
decade of research has revealed further depths of complexity. The discovery
of ever more intricate structures and systems with each increase in
knowledge—including vastly complex DNA topologies and a vast and growing
inventory of mini-RNA regulator molecules—tells us there is probably much
more to uncover. What we glimpse now may be only a tiny fraction of what
remains to be discovered.
[Denton, Miracle, p16]
“As Erica Hayden confessed in the journal Nature,
“As sequencing and other new technologies spew forth data,” the complexity
unearthed by cell biology “has seemed to grow by orders of magnitude.
Delving into it has been like zooming into a Mandelbrot set… that
reveals ever more intricate patterns as one peers closer at its boundary.””
[Denton, Miracle, p16]
1B: Maintain a shared ‘clock’…
In unicellular organisms, the internal sub-parts have
to be synchronized for the intermediates to be taken up into the next pathway.
The synchronization task is critical, and of course, it is much worse for
multicellular life…
The mechanisms used in timing are amazing, having to
overcome both randomness and unexpected change…
And—as with most ‘seemingly intelligent cellular
processes’—they are ‘poorly understood’
… LOL
“The correct timing of molecular
and cellular events is critical for embryo development, cell/tissue
homeostasis, and to functions in all organisms
throughout their whole lives. Thus, it plays a major role in
biology. Despite this obvious key role of timing in all biological processes, we
do not know exactly how cells and organisms measure time and how they translate
the information on time flow to the correct regulation of molecular processes…
Cells and organisms also measure time flow, react to time passing, and may
modify processes using time through accelerating, delaying, or postponing
certain reactions … Biochemical processes must progress using time with
appropriate timing, and the correct sequence of events is strictly
controlled using time to ensure their full coordination and purposefulness
for the cell, tissue, and organism’s life. Checkpoint mechanisms monitor if
the necessary processes have been completed before starting new ones. Thus, the
precise timely coordination between molecular events/reactions/pathways and
their specific regulation in different conditions allows for the harmonious
functioning of cells, tissues, organs, and whole organisms .. The review article by Yool Lee and Jonathan P.Wisor
discusses how circadian clocks reciprocally interact with other signaling
and metabolic factors to coordinate daily rhythms. Numerous cellular and
animal models demonstrate the presence of functional circadian oscillators at
multiple levels, ranging from individual cells, like neurons or fibroblasts, to
the brain and peripheral organs. These oscillators are tightly coupled
to the timely modulation of cellular and bodily responses to physiological
and metabolic cues.”
[Kubiak, J.Z.; Kloc, M. “How
the Timing of Biological Processes Is Controlled and Modified at the Molecular
and Cellular Level” 2.0. Biology 2024, 13, 170.]
“Many developmental processes depend
on precise temporal control of gene expression.. This timing mechanism acts across a
cell division that occurs in the neuroblast lineage and is influenced by the
asymmetry of the division. Finally, we show that positive feedback of receptor
expression through the canonical Wnt pathway enhances temporal precision. We
conclude that robust cell-intrinsic timing can be achieved by combining
regulation and feedback of the timekeeper gene….
To measure time, biological clocks
generally utilize a component that increases or
decreases in activity or abundance (a timer) or cycles through a high and low
state (an oscillator) and triggers a response when a
specific threshold is crossed (--). …. In clocks that act across multiple cells, robustness
can be increased through averaging, synchronization through intercellular
communication (--) or entrainment using external signals (--). The cell
intrinsic mechanisms that control the timekeeping mechanism itself are,
however, still poorly understood… [Journal Editor's evaluation: This paper (above) deals with an important unsolved
problem in developmental biology: how cells execute their dynamics at the right time. The
study combines compelling quantitative single-cell and single-transcript experiments
with genetic perturbations and computational modelling and provides important insights into how the timing of
transcription is regulated.]
[Erik
S Schild, Shivam Gupta, Clément Dubois, Euclides E Fernandes Póvoa, Marie-Anne
Félix, Andrew Mugler, Hendrik C Korswagen (2023) “Precise temporal control of
neuroblast migration through combined regulation and feedback of a Wnt receptor”, eLife 2023;12:e82675]
“Timing
is essential for many cellular processes, from cellular responses to external
stimuli to the cell cycle and circadian clocks. Many of these processes are
based on gene expression. For example, an activated gene may be required to reach in a precise time a
threshold level of expression that triggers a specific downstream process.
However, gene expression is subject to stochastic fluctuations, naturally
inducing an uncertainty in this threshold-crossing time with potential
consequences on biological functions and phenotypes. Here, we consider such
timing fluctuations, and we ask how they can be controlled. Our analytical
estimates and simulations show that, for an induced gene, timing variability
is minimal if the threshold level of expression is approximately half of
the steady-state level. Timing fluctuations can be reduced by increasing
the transcription rate, while they are insensitive to the translation rate. In
presence of self-regulatory strategies, we show that self-repression reduces
timing noise for threshold levels that have to be reached quickly, while
self-activation is optimal at long times.”
[“Stochastic timing in gene expression for simple regulatory
strategies”. Alma Dal Co1, Marco Cosentino Lagomarsino2,3,4,
Michele Caselle1 and Matteo Osella1, Nucleic Acids Research, 2017, Vol. 45, No.
3 1069–1078]
“Circadian
rhythms are observed in a wide variety of biological, physiological and
metabolic processes in most organisms. These rhythmic cycles are
governed by an endogenous circadian clock. Thus, a crucial function of
the endogenous circadian system would be to anticipate and adapt to environmental
changes in light, temperature, food and even mate availability, and organize
behavior and physiology to these changing situations. In addition, the
circadian system plays a regulatory role by the temporal coordination of
physiological, cellular and molecular processes such that synergistic processes are timed to coincide, whereas
processes that are conflicting are temporally separated. There is
increasing evidence that a smoothly running endogenous clock is crucial for
energy balance in organisms from cyanobacteria to mammals [--]. … The
rhythm can also be entrained by changes in timing of food and temperature
(although the period of rhythm can be stable over a wide range of temperature—i.e.,
it can be temperature compensated) [--]. The importance of the circadian
clock for organismal homeostasis was demonstrated by loss of sleep/wake cycles
and metabolic defects following targeted disruption of core clock genes in mice
[--]. Additional roles of clocks in basic
cellular pathways have also been elucidated such as regulation of nutrient
and energy balance, cell cycle, DNA-damage repair and xenobiotic detoxification
[---]. In view of these data, it should not come as a surprise that disruption
of circadian rhythms would increase vulnerability to stressors, accelerate
aging and could lead to various pathologies including cancer …. Circadian
rhythms have been described in both prokaryotes and eukaryotes, and many
molecular features associated with the circadian clock are evolutionarily
conserved.”
[“Nature’s
Timepiece—Molecular Coordination of Metabolism and Its Impact on Aging”. Andrea
Bednářová, Dalibor Kodrík and Natraj Krishnan, Int. J. Mol. Sci. 2013,
14, 3026-3049].
"The
molecular clock network consists of transcriptional and translational feedback
loops where core clock genes are transcribed
and these translational products inhibit their own transcription. These timed
transcriptional and translational feedback processes generate the
molecular rhythms, which eventually translate to the circadian rhythms
observed in most organisms. The basic organization of the circadian clock
has a hierarchy—with every cell in the organism having an
autonomous clock, and different cells and
tissues are synchronized to oscillate with the same phase. This
orchestration occurs via the “central” or “master” clock, which is entrained by
input from zeitgebers. Genetic analyses have revealed numerous clock genes in different species and most
studies to date have focused on understanding the molecular mechanisms of the
rhythms generated by these core clock genes.”
[“Nature’s Timepiece—Molecular Coordination of
Metabolism and Its Impact on Aging”. Andrea Bednářová, Dalibor Kodrík and
Natraj Krishnan, Int. J. Mol. Sci. 2013, 14, 3026-3049].
“The inherent probabilistic nature of biochemical
reactions and low copy numbers of molecules
involved result in significant random fluctuations (noise) in protein
levels inside isogenic cells inhabiting the same environment (--). Although the
origins of stochastic gene expression have been extensively studied across
organisms, the impacts of the noisy expression of key regulatory proteins on
the timing of intracellular events is underappreciated (--). Identifying
regulatory motifs that buffer randomness in the timing of intracellular
events has important consequences for disparate cellular processes, such as
apoptosis, cell-cycle control, cell differentiation, and sporulation, where
precision required for proper system functioning depends on regulatory
molecules reaching critical threshold levels at the right time… In another
example, proper timing of yeast cell division is ensured by the precise
expression of a regulatory protein, Cln, up to a critical threshold level (--).
Despite these significant impacts, how cells
maintain precision in event timing despite noisy gene expression remains poorly understood. … In summary, our
study uncovers mechanisms for generating precision in the timing of
cellular events given the unavoidable constraints of stochastic gene expression
and dilution from cellular growth.”
[“Optimum Threshold Minimizes Noise in Timing of
Intracellular Events”. Kannoly et al., iScience 23,, 101186, June 26,
2020 ]
Here is a table/list of the biological clocks in use
by cells – notice the complexity that this implies:
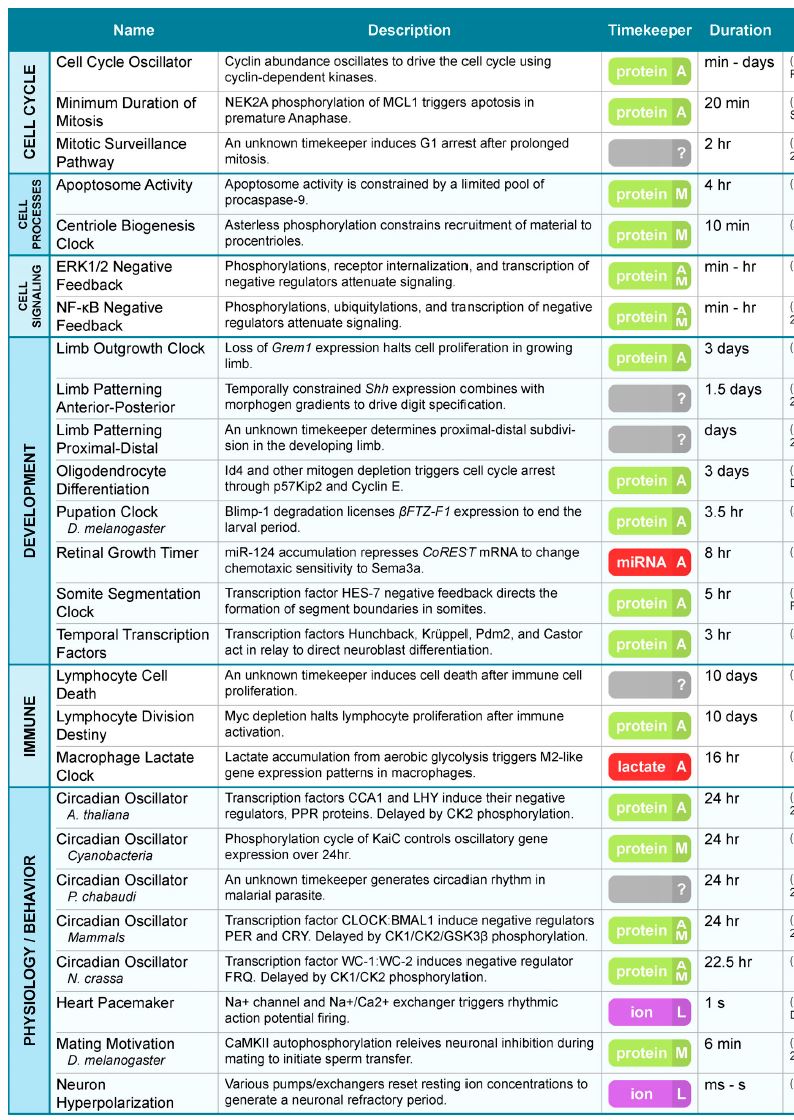
From “Keeping track of time: The fundamentals of
cellular clocks”, Colin R. Gliech and Andrew J. Holland, J. Cell Biol.
2020 Vol. 219 No. 11, page3]
…………………..
Timing is important to spacing
accuracy, as seen in size-control for example:
“Growth of a cell and its subsequent division into
daughters is a fundamental aspect of all cellular living systems. During these
processes, how do individual cells correct size aberrations so that they do
not grow abnormally large or small? How do cells ensure that the
concentration of essential gene products are maintained at desired levels, in
spite of dynamic/stochastic changes in cell size during growth and division?
Both these questions have fascinated researchers for over a century. We review
how advances in single-cell technologies and measurements are providing unique
insights into these questions across organisms from prokaryotes to human cells.
More specifically, diverse strategies based on
timing of cell-cycle events, regulating growth, and number of
daughters are employed to maintain cell size homeostasis. ….”
[“Cell size control and gene expression homeostasis in
single-cells”. Cesar A. Vargas–Garcia1, Khem Raj Ghusinga, and Abhyudai Singh. Curr
Opin Syst Biol. 2018 April ; 8: 109–116]
“All living systems display remarkable spatial and
temporal precision, despite operating in intrinsically fluctuating environments.
It is even more surprising given that biological phenomena are regulated by
multiple chemical reactions that are also random. Although the underlying
molecular mechanisms of surprisingly high precision in biology remain not well understood, a novel theoretical
picture that relies on the coupling of relevant stochastic processes has
recently been proposed and applied to explain different phenomena … In this
review, we provided novel microscopic insights into how biological systems
achieve precise control over spatial and temporal processes despite the
inherent randomness of molecular interactions. By examining bacterial
cell-size regulation and the timing of cell lysis in l bacteriophage, we
demonstrate that cells can harness stochastic
processes to ensure reliable outcomes. In bacterial cells, balanced
growth and division rates lead to stable size distributions, whereas, in l
phage, holin protein accumulation triggers timely lysis, achieving effective
threshold-like behavior. These studies highlight universal
mechanisms by which cells adapt precision strategies in size and timing,
advancing our understanding of cellular regulation.”
[“Microscopic origin of the spatial and temporal
precision in biological systems”. Anupam Mondal and Anatoly B. Kolomeisky. Biophysical
Reports 5, 100197, March 12, 2025
Although we
are mostly focused on unicellular organisms here, most of it applies to
multi-cellular life also:
“Multi-cellularity would be impossible without communication, as the existence
of the multicellular state hinges upon making sure that the cells’ decisions are perfectly synchronized, and that cells do not act independently of others. “ [Adami, p. 509]
“Genetic programs did not begin with animal cells.
Indeed, all prokaryotes and even viruses have genetic programs. But animals
stand apart for their genetic programs because most of them—the ones
we are interested in here are associated with the emergence of different cells—run
sequentially, one after another, and in parallel in the different cells that
make up the organism. As I have said, the core of these programs is the
sequence of events created by the chemistry inside the cell, but then something needs to coordinate these across cell
populations and, moreover, across different cell populations within the
organism. We do not yet know how this
happens, but we do know that the
cell is responsible for it and that the signaling kit that we can see for the
first time in sponges has much to do with it. Once again, genes and their
chemical actions are under the control of cells.” [Arias, Master, p. 115]
“From this perspective, the genes in a cell are
functionally similar to the different agencies in a self-maintaining
ecosystem. The reactions they trigger must be coordinated or
mutually adapted so that the one produces what the others consume, and
the system as a whole remains in balance—while being quick to adjust to
fluctuations in the concentrations of the different molecular
"species," or to the entry of food molecules that must be consumed or
toxins that must be neutralized. Thus, for a cell to survive, its genes must efficiently cooperate, forming a
synergetic whole where everything needed by one process is produced by one
or more other processes, and vice versa. Note that these processes
generally include reactions that transport resources into the cell and waste
out of the cell. Therefore, the system is not closed in the thermodynamic
sense, only in the organizational sense.” [Purpose, p. 95]
…………………………………………………………………………………………….
1C. Respond to internal
threats (e.g. DNA and cellular repair mechanisms)…
We noted above that the cell exerts
many control forces over metabolic processes (esp. relating to timing).
The complexity of the cell and its
systems present a large ‘attack surface’ for deleterious impacts. The
management system of the cell and the information center (DNA) of the cell, are
BOTH fallible, and if compromised enough, can trigger the failure of the cell.
These management capabilities are
therefore essential and must be protected (and repaired when damaged) at all
costs.
Everything breaks, and everything
in the cell requires monitoring and repairing and --sometimes—destroying and
disposing.
Barbara McClintock received the
Nobel Prize in 1983, for her work in
genetic change. She had this to say at the award ceremony:
“The conclusion
seems inescapable that cells are able to sense the
presence in their nuclei of ruptured ends of chromosomes and then to
activate a mechanism that will bring together and then unite these ends,
one with another ... The ability of a cell to sense these broken ends, to direct them toward each
other, and then to unite them so that the union of the two DNA strands is
correctly oriented, is a particularly revealing example of the sensitivity
of cells to all that is going on within them... … We know nothing, however, about how the cell senses danger and instigates
responses to it that often are truly remarkable.” [given in Davies, Demon,
pp. 127-128]
The cell does this through
diverse and complex repair mechanisms, that vary by organism.
The events of damage fall into three
categories:
1.
Genetic
damage – DNA focused, damage to the information center
2.
Phenotypic
mutations –
a.
errors
in creation of mRNA (transcription)
and then
b.
errors
in making polypeptides (translation),
and then
c.
errors
in making working proteins (folding)
3.
Non-genetic
cell elements (membrane, mitochondria, etc)
·
The
cell concentrates on #1, Genetic damage , since the DNA contains much of
the information needed for operations and it is the only element that is sure
to be passed down to the next generation.
·
Errors
in #2, Phenotypic mutations , are less severe (but much more common),
since they do NOT typically pass down to the next generation, they rapidly
degrade, and they can easily/quickly be replaced.
·
Some
damages in #3 can be fatal, so they have mechanisms to repair them too.
Complex organisms could not
function without these repair functions – the error rates are too high:
“With a natural
error rate in DNA copying, complex organisms could not function. It would
produce hundreds of thousands of errors that would destroy the organism.
Furthermore, the error-correcting process needs to be able to identify and
target the errors. The error-correcting process cannot function without
this ability. The double helix structure enables the fitness of two nucleotides
to be detected. Without this, an error could not be detected. But the double
helix, in itself, does not explain how the corrections happen. … When cells divide, DNA is replicated with
amazing accuracy. But this does not occur by accident. It occurs through an
equally amazing cellular process. DNA does not achieve this by itself. On
its own, there would be hundreds of thousands of mistakes every time the genome
is copied. Accurate replication can only be performed in a cell with all the cellular error-correcting machinery to
reduce an error rate of 1 in 10,000 to just 1 in 10 billion. The
correcting process involves an army of unique proteins olymeriz by the cell.
Furthermore, this provides a way for the cell to alter the DNA, because it is a
targeted process.” [Living Systems, pp. 16ff]
“What if they
could actively manipulate their own genomes? Actually, it
is clear that they do. Sexual reproduction involves several
slicing-and-dicing genomic reconfigurations, some random, some supervised. Many
intermingling methods are out there, each of which involves cells shuffling their own DNA in a carefully
arranged manner. And sex isn’t the only example. Correcting
the errors that occur during DNA replication requires another set of
genomic management operations. Most of the primary damage to DNA, for example
by radiation or thermal disruption, never
makes it to the daughter cells because it is repaired first.
Human DNA would suffer devastating mutational damage, estimated at an
overall 1 per cent copying error rate per generation, without all the
in-house, high-tech proofreading, editing and error correcting which reduces
the net mutation rate to an incredible one in 10 billion. So cells are able
to monitor and actively edit their own genomes to a high degree of fidelity in
an attempt to maintain the status quo.” [Davies, Demon, pp. 127f]
Because of the all-important DNA, the
cell devotes a ton of resources to it!
“Evolution of life
is fully based on digital information transmission processes —across
generations via genome replication and from the genome to the effector
molecules (RNA and proteins). No information transmission channel is
error-free, as first formally asserted by Claude Shannon, who founded
information theory by connecting information transmission with the laws of
thermodynamics. As noted in Chapter 2, the fidelity
of genome replication cannot be lower than a certain minimum;
conversely, the mutation rate cannot exceed a certain threshold, to avoid the
mutational meltdown of the population. .. Selection for a sufficient
fidelity of replication (and more generally all processes of information
transmission) is one of the central aspects in all evolution. This is immediately obvious from the enormous diversity,
complexity, and multilayer organization of repair systems that exist in all
cellular life forms (---). In prokaryotes, up to 10% of the coding capacity of the
genomes may be dedicated to repair system components that act at all
stages of DNA replication and also eliminate various mutational lesions that
occur outside the replication process.” [Koonin, pp. 274-276]
The repair
mechanisms for DNA are very complex, and must
have been present in the first cells, since – without their
repair work—the replication error rate would have doomed the downstream
progeny.
“Even when
selection and adaptation clearly are involved, the manifestation of these
factors of evolution often (possibly, most of the time) is quite
different from the (neo) Darwinian idea of “improvement.” Often adaptations have to do with maintaining the
integrity of cellular organization, preventing malfunction, and performing
damage control. In a sense, this is a trivial statement of fact, considering
the expanse and complexity of the molecular machinery that is dedicated to
quality control of each of the major information transfer processes: Systems
of DNA repair and protein degradation, and molecular chaperones are all
cases in point. Moreover, much, if not most, of the evolution of protein-coding
genes appears to be driven by selection for robustness to misfolding. In
multicellular organisms, the importance of the selection for prevention of
malfunction is apparent at the level of cell and tissue interactions, as
illustrated by the highly complex systems of programmed cell death. In
retrospect, all these findings may appear quite intuitive, considering how advanced,
complex, and, in a variety of ways, optimized cells and even individual protein
or RNA molecules are. Once these complex systems are in place—and evolutionary reconstructions clearly show that they
have been in place for most of the history of life, that is, more than 3.5
billion years—quality control and damage prevention indeed become the bulk of
the “work” of evolution, the importance of occasional new
adaptations notwithstanding. This realization places an enormous burden on the
early, precellular stages of evolution when change must have been rapid and the
roles of positive selection along with constructive neutral evolution must
have been much greater than they were during the subsequent 3.5 billion
years of evolution. In a sense, almost everything “really interesting” in the
evolution of life occurred during its relatively brief, earliest stages
antedating the “crystallization” of the basic cellular organization. Certainly,
major exceptions exist, such as the emergence of eukaryotic cells or multicellular
eukaryotic organisms, but there is no doubt that most of the fundamental
evolutionary innovations are crammed into the earliest 5% of the history of
life.” [Koonin, pp. 401-402]
There are multiple types of damage
that can occur, but these two diagrams will show both the typical sources of
mutation, and the complexity of a few of the repair mechanisms:
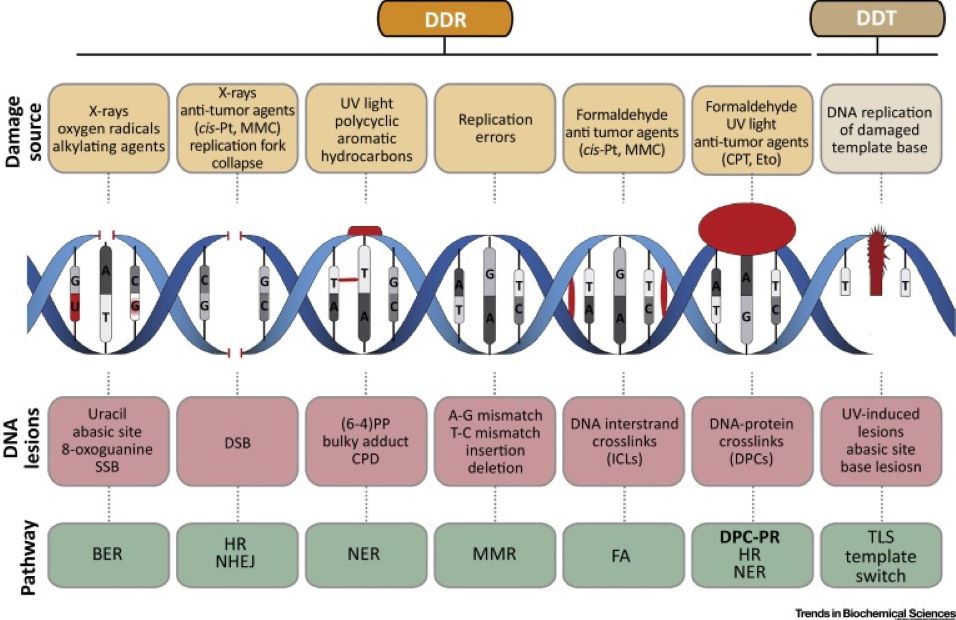
There are more repair mechanisms than
in the below image, but this diagram illustrates the pathway complexity (but
only at a high level!) of a few of the repair mechanisms:
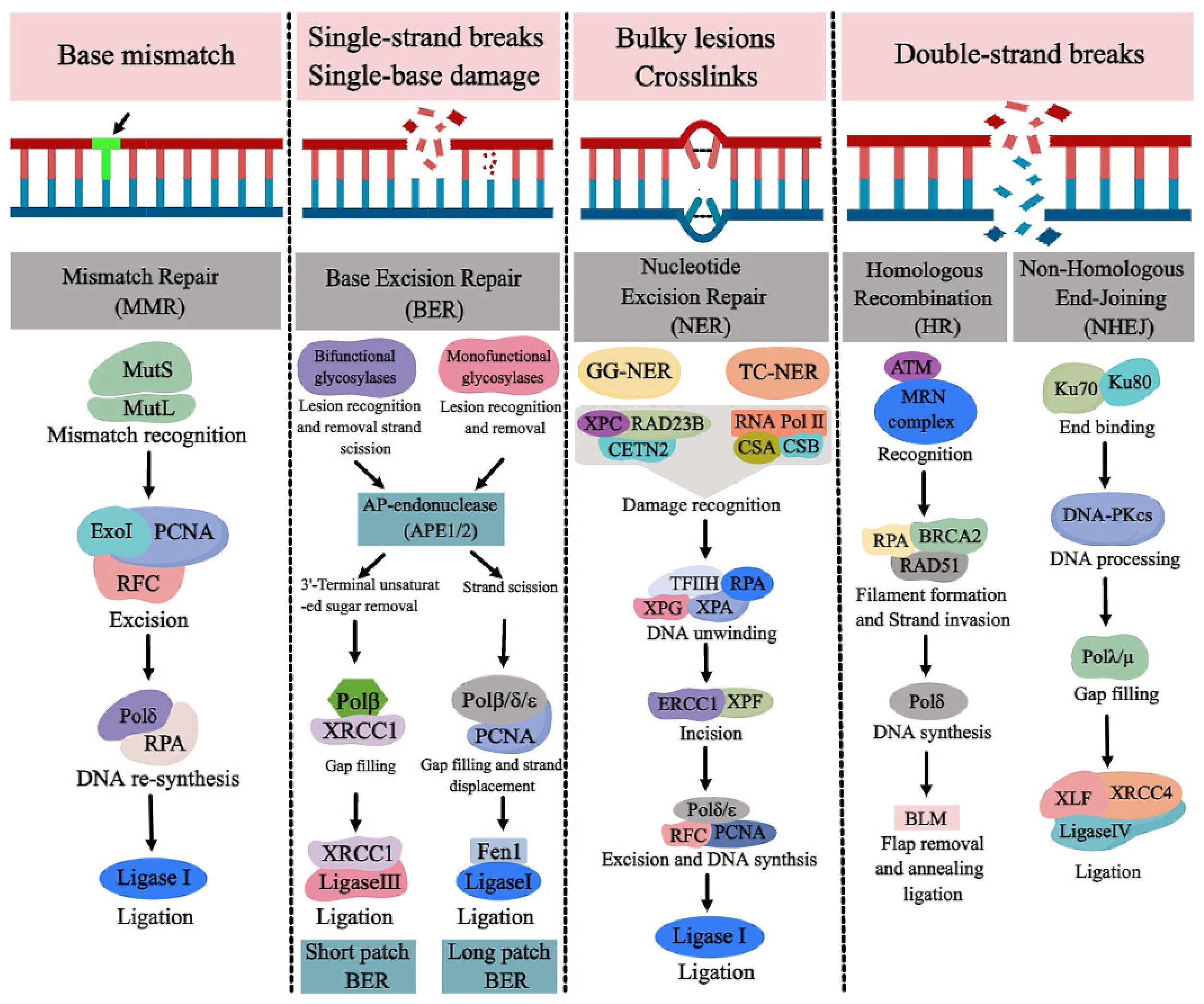
These are fast, dedicated pathways
– the Mismatch Repair (MMR) above in the left column uses between 200 and 300 total molecules
(typical repair), involving between 100,000 and
150,000 atoms in those molecules… for this one specific function.
Replication fidelity is fairly
small, but the repair mechanisms powerfully reduce errors:
·
DNA
polymerase alone: ~10⁻⁴ to 10⁻⁵ (1
error per 10,000-100,000 bases)
·
+ 3’ to 5’
exonuclease proofreading:
~10⁻⁶ to 10⁻⁷ (100-1,000x improvement)
·
Mismatch
repair (MMR): ~10⁻⁹ to 10⁻¹⁰ (another
100-1,000x improvement)
The error rates of phenotype
mutations go UP as the process gets farther away from the DNA:
“The error rate
of transcription is much higher than that of replication, and the error rate of translation is
much higher yet (see Figure 9-4). Although the experimental measurements of
the amino acid misincorporation rate during translation are scarce and
limited to a few model systems, it is clear that
the fidelity of translation is almost shockingly low. Indeed, the rate
of non-cognate amino acid incorporation is 10(-4) to 10(-5)—interestingly,
close to the replication error rate in RNA viruses. Thus, about 20% of the
protein molecules synthesized in any cell contain at least one wrong amino acid
(---). The consequences of errors of transcription and translation,
sometimes aptly called phenotypic mutations, obviously are
less dramatic than the consequences of genetic mutations, for the very reason
that phenotypic mutations are generally not inherited (notable
exceptions exist, such as reverse transcription followed by the
incorporation of a DNA copy of a mistranscribed RNA into the genome, ---). Given
the relatively short lifetime of any RNA or protein molecule, no phenotypic
mutation can have a major fitness effect on its own, so it is not surprising
that much greater error rates are tolerated for phenotypic mutations
than for genetic mutations. However, it is equally obvious that excessively
high rates of phenotypic mutation are incompatible with life. Thus, as is the
case with DNA repair systems, multiple mechanisms for keeping transcription
and translation errors in check certainly exist. Proofreading activity of
DNA-dependent RNA polymerases has been detected and shown to decrease the
error rate by orders of magnitude (---). Moreover, still poorly characterized processes of
post-transcriptional repair of methylation damage in RNA have been discovered
as well (---). Probably the best understood of the mechanisms that control the
rate of phenotypic mutations is proofreading
by aaRS, in which aminoacyl-tRNAs charged with non-cognate amino
acids are hydrolyzed and recycled (---). The aaRS proofreading is
complemented by the downstream ribosomal proofreading, in which the
ribosome rejects non-cognate tRNAs (---). However, a large increase of the
translation accuracy seems to clash with the requirement of a high rate of
protein production. A substantially increased translation fidelity can be
readily achieved by mutating specific positions in rRNA or ribosomal proteins,
but these mutations are deleterious, apparently, because of slow translation (--).
“
[Koonin, pp. 277ff, and diagram below.]
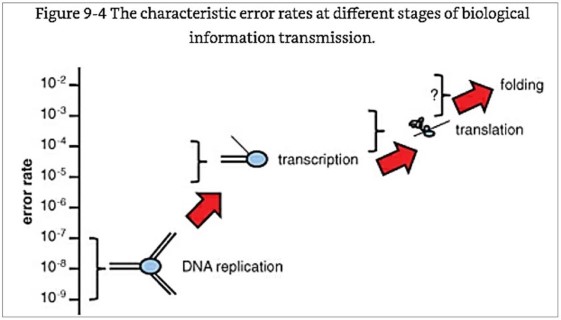
The most common
type of error is in the complex protein folding step—where
the final shape of the polypeptide is important in becoming a useful protein.
From Koonin’s explanation [Koonin, 277-279]:
“The principal deleterious effect of mistranslation
is thought to be protein misfolding (---), although amino acid misincorporation
corporation at catalytic sites certainly could be an additional factor. … Regardless,
although protein folding is not usually viewed as an information-transmission
process, this is what it actually is. Indeed, folding involves the flow of
information from the one-dimensional amino acid sequence to the
three-dimensional protein structure. Exactly the same applies to structural
RNAs. The rate of misfolding is hard to determine experimentally, and this has
not been done for large sets of proteins or RNAs. If the trend of error rates
depicted in Figure 9-4—the farther from the genome, the less accurate an
information transmission step is—is any indication, the error rate of
folding is expected to be even greater than the error rate of translation. This
prediction also stands to reason, given the enormous complexity of the
folding process and the vast number of (mis)folding pathways that are, in
principle, available to a folding protein or RNA molecule (---). Given the high
complexity of the folding landscape, the seminal discovery (made originally by
Christian Anfinsen and subsequently confirmed in numerous experiments) that
proteins can spontaneously fold into the native conformation came as a
considerable surprise.
“Almost 50 years after Anfinsen’s discovery, it
remains a matter of debate whether spontaneously folding proteins find the
global or a local free energy minimum. What has become clear is that only
small proteins fold spontaneously; the majority
of proteins require special molecular devices, namely other proteins known as
chaperones, to fold into the native structure. Chaperones
function in a remarkable manner: The chaperone molecules form a “cage” (known
also as the Anfinsen cage) that isolates the folding protein from the
cytoplasm and partially unfolds it, thus facilitating the search for the
native conformation (---). Most of the chaperones are abundant, highly
conserved, high-status proteins. Originally, some of the chaperones were
discovered as “heat shock proteins;” that is, proteins that are strongly
upregulated at elevated temperature (and, as shown later, other stress
conditions) and counteract protein misfolding, which is enhanced by the stress
(---). Although not as well characterized,
protein chaperones mediate RNA folding as well (---). On the whole, the control over protein (and probably RNA)
folding undoubtedly is a major function in all cells.
“Beyond the
chaperone-type devices, all cells dedicate a
versatile repertoire of molecular machines to controlled degradation of
proteins, particularly misfolded ones, and RNA. Similarly to
molecular chaperones, these machines— the proteasome, in the case of
proteins and the exosome (degradosome in bacteria), in the case of
RNA—are ubiquitous in the three domains of life, highly
abundant in most cells, and subject to regulation under stress (---).
Furthermore, these machines, along with additional back-up systems of regulated
proteolysis, are major intracellular consumers of energy (ATP). Bacteria
additionally possess a highly conserved system of the so-called
trans-translation that releases stalled ribosomes from aberrant mRNAs on
which translation fails to terminate properly and targets such mRNAs and
their (also aberrant) protein products for degradation (---).”
Beyond these gene-based
issues, we have repair mechanisms for the other parts of a cell:
1.
Organelle-Level
Repair
·
1.1
Mitochondrial Quality Control
(eukaryotes only)
o Mitochondrial DNA
repair
o Mitochondrial unfolded
protein response
o Fusion and fission
dynamics
o Mitophagy (selective
autophagy)
·
1.2
Other Organelle Maintenance
o
ER-phagy
(ER turnover)
o
Pexophagy
(peroxisome turnover)
o
Lysosomal
repair
2. Bulk Degradation and
Recycling
·
2.1
Autophagy
(eukaryotes only, although maybe broader)
o
Macroautophagy
o
Microautophagy
o
Chaperone-mediated
autophagy
o
Selective
autophagy (mitophagy, pexophagy, etc.)
3. Membrane Repair
·
Plasma
membrane repair
·
Organelle
membrane repair
·
Lipid
remodeling and replacement
4. Oxidative Damage
Response
·
ROS
neutralization (antioxidant enzymes)
·
Lipid
peroxidation repair
·
Protein
oxidation reversal
1D: Reproduce (when it senses that
conditions are right for that)….
At a simplistic
level, cells reproduce by making ‘two cells within one cell enclosure’ and then
splitting this in half – with two identical cells, now separated.
Literally,
everything inside the cell is DUPLICATED and/or elongated. The two sets of
things move away from each other (in the elongated cell envelope) and then the
cell is cut in half.
You have probably
seen diagrams of this in high-school biology class, something like this:
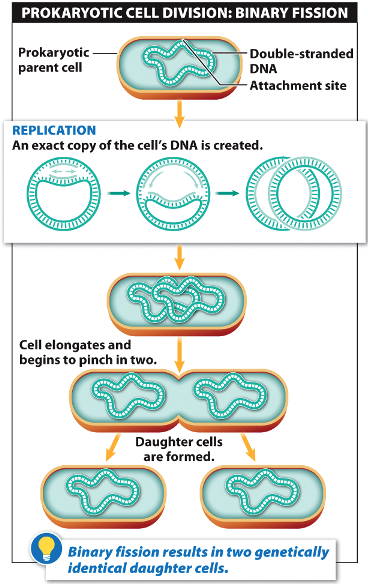
It is called
‘binary fission’ (or in some cases, ‘mitosis’). It is a super complex
operation, but can occur in prokaryotes every 10-30 minutes, and in humans, we
have 25 MILLION cell divisions PER SECOND.
We know very
little about how this is controlled, orchestrated, or even initiated – at the
agent level. So, Arias, [Master, pp. 79ff] on eukaryotes:
“There is much
we still don’t know about mitosis, particularly
about the control and precise engineering of the spindle, centrioles, and
chromosomes. But we can say that the answer to many of these
questions cannot lie in the genes. Throughout the process of mitosis, genes
remain inert, passive molecular structures. The chromosomes, on which genes
reside, are copied and transported but have very limited involvement in what’s
happening. Instead, it’s the organelles within the parent cell that sense
space, balance forces, partition, and move chromosomes around. For example,
we know that the timing of mitosis is not decided by one particular gene or
even a collection of genes. Whether a cell divides or not relies on
the assessment of many of its features: its volume, composition, stiffness,
availability of nutrients, and signals from its
neighbors. None of these can be directly linked to genes, though
they will have an impact on the activity of the genome by determining the
expression of certain genes whose products might be needed for a particular
event—but the cell will decide what happens. “
“We are starting to learn how the cell does this; for
the moment, however, we only have glimpses of
the process. For example, a protein called mTOR plays a
central role in gathering the information outlined above, determining
when a cell will divide by controlling its growth. Found in all cells in
the body, mTOR senses the cell’s health and available nutrients and,
based on this information, decides whether it
should make more nutrients, divide, or die. In a situation of
stress, mTOR is able to read the state of the cell integrating many variables,
measure them against the available nutrients, and, if possible, promote
biosynthetic metabolism and growth. We still do not understand how it makes
the decision.”
“In one dramatic
instance, mTOR controls a process called autophagy, meaning self-devouring,
in which, when stressed or running out of resources, a cell turns on itself and
recycles its proteins and organelles, usually damaged ones, as a source of
energy to stay alive. mTOR is a gatekeeper of autophagy. Usually it suppresses
this process, favoring less dramatic ways to create mass and power, but if
it assesses that there is not enough metabolic substrate for this, it will be
switched off, allowing autophagy to proceed as an emergency system. This is
an example of a vital activity that happens far from the realm of genes. Of
course, mTOR is encoded by a gene, but this gene only provides the instructions
for assembling the protein. It is mTOR that does the heavy lifting that keeps
the cell alive. The protein’s function is determined not by the activity of the
gene but, as with any sensor, by the protein’s interactions with other proteins
and the wider chemical environment.”
And—in keeping
with the data mentioned (profusely…sigh) above, this process is monitored
CLOSELY, and even PAUSED if problems are sensed:
“Complex cells
with a nucleus (eukaryotes) display a tightly controlled multistage cell
division cycle. Each stage involves intricate processes such as cell
growth, DNA replication, and accurate transmission of genome copies to daughter
cells. Elaborate biochemical reactions regulate passage from one stage
to the next. On top of the stage-to-stage control circuitry, a
self-monitoring system makes sure everything comes out right. If the
different biochemical and biomechanical processes fall out of sync, or if there
is either a mistake or damage, sensory molecules
detect the problem. They activate a checkpoint to hold up the entire cycle
until everything has been set right for renewed progress. Cells set distinct
checkpoint systems for growth and division.” [Shapiro, p. 198]
1E: Respond to External events
(especially threats) as a unit, and …
This is one of
fundamental characteristics of life – and ONLY of life and ALL of LIFE.
“Moreover, modern
organisms react to environmental signals and changes—this reactivity is one of the hallmarks of life, and
importantly, the ability to learn how to react to such signals also requires
explanation in order to fully solve the origin of life puzzle.” [Prebiotic,
p. 41]
“All living cells sense
and respond to changes in external or internal conditions. Without that
cognitive capacity, they could not obtain nutrition essential for growth,
survive inevitable ecological changes, or correct accidents in the complex
processes of reproduction. Wherever examined, even the smallest living cells
(prokaryotes) display sophisticated regulatory networks establishing
appropriate adaptations to stress conditions that maximize the probability of
survival. Supposedly “simple” prokaryotic organisms also display remarkable
capabilities for intercellular signaling and multicellular coordination. These
observations indicate that all living cells are cognitive.” [Shapiro, p. 928]
Bacteria, for
example, have the ability to sense these external factors—through their cell
wall—and take actions based on them. Here is a list of such sensors and what
they do:
|
NUTRIENTS
(Attractants) |
Examples |
Cellular Response |
|
|
Amino acids |
Serine,
aspartate |
Smooth
swimming toward source; continued flagellar rotation |
|
|
Sugars |
Glucose, galactose, ribose, maltose |
Smooth
swimming toward source; uptake system activation |
|
|
Dipeptides |
Various
dipeptides |
Smooth
swimming toward source |
|
|
Oxygen |
O₂ |
Aerotaxis toward optimal O₂ levels |
|
|
TOXINS/REPELLENTS |
Examples |
Cellular Response |
|
|
Heavy metals |
Nickel
(Ni²⁺), cobalt (Co²⁺) |
Tumbling/direction change (away from source) |
|
|
Weak acids |
Various
organic acids |
Tumbling/direction change (away from source) |
|
|
Toxic compounds |
Leucine, indole |
Tumbling/direction change (away from source) |
|
|
PHYSICAL PARAMETERS |
Examples |
Cellular Response |
|
|
pH changes |
Acidic/alkaline conditions |
Gene
expression changes; stress response proteins; motility adjustments |
|
|
Osmolarity |
High/low salt concentration |
Porin
expression regulation; compatible solute synthesis |
|
|
Temperature |
Heat/cold stress |
Heat
shock proteins; cold shock proteins; membrane fluidity changes |
|
|
QUORUM SENSING |
Examples |
Cellular Response |
|
|
Population density signals (Gram-negative) |
Acyl-homoserine lactones (AHLs) |
Biofilm
formation; virulence factor production; bioluminescence |
|
|
Population density signals (Gram-positive) |
Oligopeptides |
Biofilm
formation; competence; sporulation; virulence |
|
|
CELL DENSITY / CONTACT |
Examples |
Cellular Response |
|
|
Cell-to-cell contact |
Physical proximity |
Conjugation; biofilm maturation; coordinated
behavior |
|
Different species
can also sense other factors, which they respond to:
“Bacteria turn out
to be highly versatile when it comes to guiding their swimming motility [---].
Among the 5 E. coli MCP receptors is one that senses oxygen and guides
“aerotaxis” to more oxygenated zones of the growth medium. In addition to
chemotaxis and aerotaxis, some bacteria like M. magnetotacticum
have receptors called “magnetosomes” that sense the Earth’s magnetic field.
Such sensing enables “magnetotaxis,” which effectively allows aquatic cells to
move either up or down in a controlled manner [--]. Many bacteria also have
light receptors that guide “phototaxis” (movement toward light), “photophobia”
(movement away from light), or more complex behaviors in the illuminated
environment [--].” [Shapiro, p. 934].
In some cases, they recognize incoming
DNA sequences as being ‘not self’!
“Bacteria have at
least three different systems [one of which is CRISPR-Cas system] for recognizing
incoming DNA and discriminating between it
and their own genomic molecules..” [Shapiro, p. 950]
[More on this in the next section
on ‘social issues’…]
1F: Occasionally MODIFY THEMSELVES as needed to
fulfill the requirements above…
This statement
would have been blasphemous up until recently, since the cell was believed to
be passive in its genetic future. Change came from mutations, and mutations
were random—the cell was only the recipient of change, and not the instrument
of change.
That old position (the
Central Dogma) is very far from the consensus view today, and runs counter to
the mass of data gathered in the last couple of decades.
“A key proposition
of the Central Dogma is that cells cannot alter DNA in a directed way.
Furthermore, there is assumed to be no facility for such alteration. However,
this proposition is not based on experimental evidence, and there is a
growing body of evidence that it is false. It is based on the idea that the
transcription process from DNA to protein cannot work backwards. But the cell
does not need to do this to alter the genome. In one sense, it merely needs to
allow changes to occur, as it can, for example, by varying the error correction
process, or by large-scale chromosome and genome rearrangements, and then
choose between the variants.……Cells can
influence the DNA, not through back-translation from protein to DNA,
but by using the processes that enable the DNA to be corrected, and precisely
this mechanism operates in the immune system. Thus, DNA is regulated by the
system, and it can also be altered by the organism.……Yet,
by reverse transcription it becomes possible to transfer sequences, including
whole domains corresponding to functional parts of proteins, from one part of
the genome to another. We know that this has happened during evolution.” [Living
Systems, p. 22]
“Organisms can
achieve genome reorganisation in several ways, not least by
splicing and translocating sections of DNA. Indeed, this process has played a
significant part in adaptive evolutionary change. It pulls apart the Central
Dogma. The genome can be reorganized. Yet such was the hold of the Central
Dogma that the evidential basis for this could not be published in
standard scientific journals for many years. The evidential basis is now substantial,
beginning notably with Barbara McClintock’s work in the 1930s, 1940s and
1950s. McClintock showed genome reorganisation in corn (maize), where large
domains are moved from one region of the genome
to another in response to environmental stress, and even from one
chromosome to another. Eventually, when it became clear that many others had
found the same process in other organisms, McClintock received the Nobel Prize
in 1983. She wrote: “In the future attention undoubtedly will be centered on
the genome, and with greater appreciation of its significance as a highly
sensitive organ of the cell, monitoring genomic activities and correcting
common errors, sensing the unusual and unexpected events, and responding
to them, often by restructuring the genome.”
(our emphasis).” [Living Systems, p. 23]
McClintock faced
‘central dogmatic’ skepticism (smile), but her experience and her
statements at the Nobel prize ceremony were incontestable [from Davies, Demon,
pp. 127-129]:
“But now we
encounter a fascinating question: can cells actively edit their genomes to change
the status quo? Decades before the work of Cairns and Rosenberg this
question was investigated in a series of remarkable experiments by the
distinguished botanist and cell biologist Barbara McClintock. Starting in her
student days in the 1920s, she experimented with maize plants and established
many of the basic properties of chromosome structure and organization we know
today, for which she subsequently received a Nobel Prize in Physiology or
Medicine — the first woman to win the prize unshared in that category. With the
help of a basic microscope, McClintock looked to see what happened to the
chromosomes of the maize plants when they were exposed to X-rays. What she
reported caused such a ballyhoo and attracted so much scepticism that in
1953 she felt moved to stop publishing her data. What was uncontentious
were her observations that chromosomes break into fragments when irradiated.
But the big surprise was the fact that the
pieces could rejoin again, often in novel arrangements.
Humpty-dumpty could be reassembled in a baroque sort of way. Chromosome
reorganization writ large might seem lethal, and it often was. But it was not
always so: in some cases, the mutant plants went on to replicate their grossly
modified genomes. Crucially, McClintock found that the large-scale mutations
were far from random; it appeared that the maize cells had a contingency plan
for the day their genomes were smashed. Even more amazingly, if
the plants were stressed, for example by infection or mechanical damage, spontaneous
chromosome breakage could occur without the benefit of X-ray disruption;
the broken ends were rejoined after the chromosome replicated. In 1948
McClintock made her most startling discovery of all. Segments of chromosomes
could be transposed — switch places on the genome — a phenomenon popularly
known as ‘jumping genes’. In the maize plants this produced a mosaic
coloured pattern.”
“Today, genomic
transpositions are recognized as widespread in evolution. It has been estimated
that up to half the human genome has undergone such genetic gymnastics.”
“On the occasion
of her Nobel Prize, awarded for ‘the discovery of mobile genetic elements’, she
had this to say:
“The conclusion
seems inescapable that cells are able to sense the presence in their
nuclei of ruptured ends of chromosomes and then to activate a mechanism
that will bring together and then unite these ends, one with another ... The
ability of a cell to sense these broken ends, to direct them toward each other,
and then to unite them so that the union of the two DNA strands is correctly
oriented, is a particularly revealing example of the sensitivity of cells to all that is going on within them... A
goal for the future would be to determine the
extent of knowledge the cell has of itself, and how it utilizes this
knowledge in a ‘thoughtful’ manner when challenged... monitoring genomic
activities and correcting common errors, sensing the unusual and unexpected
events, and responding to them, often by restructuring the genome. We know
about the components of genomes that could be made available for such
restructuring. We know nothing, however, about how
the cell senses danger and instigates responses to it that often are truly
remarkable.”
.
But the data is
too extensive at this point – cells have the ability to adjust themselves, and
the ability to sense – and time –when/what is needed.
“Recent progress
in microbiology has shown that an overwhelming
majority of DNA changes in the genome are the product of internal regulatory
and control networks, not random mutations and incremental “additive”
selection. There is also much evidence of various biases in
mutational processes (---). Indeed, rapid genome alteration and restructuring
can be achieved by a variety of mobile DNA “modules”—transposons (---),
integrons, CRISPRs, retroposons, variable antigen determinants, and more (---).
It is now also apparent that individual cells have a variety of internal
regulatory and control capabilities that can significantly influence cell
development and the phenotype. More significant, they may even provide
feedback that modifies the genome and
affects subsequent generations (---). Particularly significant are the
discoveries related to the influence of exosomes, which resemble Darwin’s
speculative ideas of pangenesis and internal migratory “gemmules” (Darwin’s
term) in reproduction, as Noble (2019) has pointed out. Exosomes also clearly
violate the so-called Weismann barrier (the assumption that genetic change is
only a one-way process). .. As Shapiro
(2011, 2) emphasizes, “The capacity of living
organisms to alter their own heredity is undeniable. Our current
ideas about evolution have to incorporate this basic fact of life.”
[Purpose,
p. 14]
And some of our
bio-engineering tools were ‘appropriated’ from processes observed in
‘primitive’ cells (e.g. CRISPR):
“Conventional
evolutionary theory teaches that targeting genome change is unnatural, a recent
human invention. Nonetheless, vertebrates have been
doing just exactly that for about 500 million years as they evolved the
intricate adaptive immune system that protects us from infections and cancer [--].
Furthermore, last year’s Nobel Prize in chemistry went to two women who
pioneered RNA-targeted genetic engineering methods based
on sophisticated systems from supposedly primitive prokaryotic cells
without nuclei [--]. Bacteria and archaea have probably been using the now
famous CRISPRs to defend themselves longer than
nucleated eukaryotic cells have existed on Earth” [Shapiro, p.38].
And the ability to
re-engineer and shift DNA shows up in so many cellular-level interactions.
Consider the responses of the ‘simpliest’ of prokaryotes to our immune
responses to their invasion:
“Although the
mammalian adaptive immune system is exceptional in the complexity and
specificity of its multiple actions on the antibody-coding loci in our genomes,
it is far from unique in using DNA manipulations to modify cell-cell
interactions. Even the simplest of
prokaryotic cells… rearrange the DNA encoding their surface proteins to alter
them so they can escape antibody-guided destruction [--]. The
general name for such genome manipulations is antigenic variation. In other
words, our immune system is in a constant NGE (natural genetic engineering) arms
race with the microbes that try to take over our bodies for their own ends.” [Shapiro, p.39].
His
conclusion—having incorporated overwhelming numbers of cases of
self-modification—is this:
“As I see it,
virtually all the major progress in using the most modern tools of genome
analysis these last 10 years favors the idea of
evolution as an active process of self-modification by organisms
endowed with a toolbox of change processes totally inconceivable at the time
DNA was identified as the molecular carrier of genetic information in 1953.”
[Shapiro, p.41].
______________________________________________________________________ _________
TWO:
When all other assemblies of
chemical structures DO NOT (!)…
As can be seen in many of the
quotes above, the ‘something different’ about life is that it ‘pursues’ something … it pursues food,
internal health, resiliency against the outside world, and even expansion of
itself further into that external world through reproduction.
Life
is always hungry…always seeking energy to persist…
“In all known cases of self-ordering, dissipative
systems that display emergent properties, autocatalysis continues until the
conditions for the system’s continuation cease. The major difference
between such nonbiological systems and biological systems is not the ability to replicate – given inputs of
matter and energy, autocatalytic sets will replicate if split into
physically separated spaces. Rather, the major difference is the ability of a biological system to “reach out and
manipulate the world on its own behalf” (Kauffinan 2000, 49). A
candle flame gutters and dies when the wax is exhausted; it doesn’t search
for more. The same is true for a Renard cell, a laser, a hurricane or a
Belousov-Zhabotinsky reaction when the conditions for their existence cease.”
[“To Be or Not To Be:
Where Is Self-Preservation in Evolutionary Theory?”. Pamela Lyon. In Transitions,
p. 109.]
“By contrast, in
conditions of nutrient limitation, a population of Bacillus subtilis will synthesize enzymes to scavenge
alternative sources of nutrients (Msadek 1999). If that fails, a
portion of the bacterial population suicides through autolysis, which means
individual cells break open their membrane and literally spill their guts,
providing a temporary source of food for the remaining members of the
population in the event conditions are about to change (--). If conditions
remain dire, the population commits to sporulation, an irreversible,
do-or-die process in which cells transform, over eight hours, into spores
capable of resisting injury by heat, chemicals and ultraviolet radiation and
remaining dormant (sometimes for years) until life-sustaining conditions
return.”
[“To Be or Not To Be:
Where Is Self-Preservation in Evolutionary Theory?”. Pamela Lyon. In Transitions,
p. 109.]
“Kauffman is correct. Many complex structures –
including networks – can and do arise spontaneously. Indeed, we can find
examples of order-for-free all around us. The problem is that such structures
do not yet have function, agency, or purpose.
They are not yet alive. Self-organization, as mathematicians and physicists use
the term, may indeed be necessary for the emergence of biological forms of
organization, but as I have argued on a number of occasions, and as Stuart
Kauffman now acknowledges, for understanding living processes it is not enough.
Despite all our efforts, the critical properties
of function, agency, and purpose continue to mark organisms (even if not
machines) apart from thunderstorms – indeed, apart from all the
emergent phenomena of nonlinear dynamical systems, remaining conspicuously
absent from the kinds of systems with which physics deals. An account of how properties of this sort might emerge from
the dynamics of effectively homogeneous systems of simple elements, however
complex the dynamics of their interaction might be, continues to elude us.
Such properties seem clearly to require an order of complexity that goes beyond
that which spontaneously emerges from complex interactions among simple
elements – a form of complexity that control engineers have been struggling to
characterize ever since the 1940s, and that Warren Weaver, Herbert Simon, and
now John Mattick and John Doyle, have dubbed organized complexity.”
[“To Be or Not To Be:
Where Is Self-Preservation in Evolutionary Theory?”. Pamela Lyon. In Transitions,
p. 109.]
Life
is always seeking something, always has a purpose
Modern biologists
have long recognized that the core difference between life and non-life was
purpose (or ‘teleology’):
“Purposefulness,
or teleology, does not exist in nonliving nature. It is universal in the living
world. It would make no sense to talk of the purposiveness or adaptation of
stars, mountains, or the laws of physics. Adaptedness of living beings is
too obvious to be overlooked.… Living beings have an internal, or natural,
teleology. Organisms, from the smallest bacterium to man, arise from similar
organisms by ordered growth and development. Their internal teleology has
accumulated in the history of their lineage. On the assumption that all
existing life is derived from one primordial ancestor, the internal teleology
of an organism is the outcome of approximately three and a half billion years
of organic evolution.… Internal teleology is not a static property of life. Its
advances and recessions can be observed, sometimes induced experimentally, and
analyzed scientifically like other biological phenomena (Dobzhansky et al.,
1977, 95–96).
“Many other
theorists over the years have expressed similar views, as Samir Okasha has
documented in his book-length study, Agents and Goals in Evolution (2018;
see also Walsh, 2015). For instance, the Nobel biologist Jacques Monod (1971,
9) concluded that “one of the most fundamental characteristics common to all
living things [is] that of being endowed with a project, or a purpose.”
Likewise, the biologist Ernst Mayr, one of the founding fathers of the
so-called modern synthesis in evolutionary biology, wrote “goal directed
behavior … is extremely widespread in the natural world; most activity
connected with migration, food-getting, courtship, ontogeny, and all phases
of reproduction is characterized by such goal orientation” (Mayr, 1988,
45).
[Purpose, p.
12]
“The huge gulf
that separates physics and biology – the realm of atoms and molecules from that
of living organisms – is unbridgeable without fundamentally new concepts. Living organisms have goals and purposes – the
product of billions of years of evolution – whereas atoms and molecules just
blindly follow physical laws. Yet somehow the one has to come out of the other.”
[Davies, Demon, pp. 1-2]
And that has even
become the ‘ground-zero premise of evolutionary biology”:
“The ground-zero
premise of evolutionary biology is that life is, in essence, a “survival
enterprise.” Living organisms are inherently contingent, dynamic phenomena
that must actively seek to survive and
reproduce. This existential problem requires that they must be
goal-directed in an immediate, proximate sense.”
[Purpose, p.
15]
“All models of
life-origin, whether Protometabolism-First or pre-RNA / RNA World early informational self-replicative
models, encounter the same dead-end: no
naturalistic mechanism exists to steer objects and events toward
eventual functionality. No insight, motive, foresight or impetus exists
to integrate physicochemical reactions into a cooperative, organized, pragmatic
effort. Inanimate nature cannot pursue the goal of homeostasis; it cannot scheme
to locally and temporarily circumvent the 2nd Law. This deadlock
affects all naturalistic models involving hypercycles, composomes and
chemotons. It precludes all spontaneous geochemical, hydrothermal, eutectic,
and photochemical scenarios. It affects the Lipid, Peptide and Zinc World
models. It pertains to Co-evolution and all other code-origin models. No
plausible hypothetical scenario exists that can convert chance and/or necessity
into an organized protometabolic scheme. “
[The First Gene,
David L. Abel, Editor 2011, pp 231-286]
Life
is working, never getting a ‘day off’ – ceaseless activity until its “time in
the sun is up”…and then it releases its resources to the next generation
of life.
1.
Within
the cell, when components are damaged beyond repair, the process of autophagy
identifies those components and ’de-commissions’ them (stopping them from
damaging others), and either recycles/reuses their internal materials (.e.g the
salvage pathways) or disposes of them to the outside world.
2.
When
enough internal processes of the cell become ineffective to maintain cell
health (e.g. homeostasis) and repair mechanisms cannot remediate them, then the
cell will schedule it own shutdown/death (apoptosis)—and it becomes then
‘just another group of chemicals’ that degrades and dissipates according to the
natural processes of chemistry and physics.
3. When cells in a
cooperative arrangement detect that the food supply is not enough to support
the groups, some volunteers sacrifice themselves to become food for the others
[example given above of Bacillus subtilis.]
All of these processes require the complex signaling and
metabolic ‘interventions’ described above.
No collection of non-living molecules does this…all the
examples of auto-catalytic systems and crystalline structures and emergent
vortices do not come close. The two are not the same….
__________________________________________________________
__________
THREE:
And when we have no real explanation of how they got here (?!) …
We have no plausible idea how it
GOT HERE (not that any understanding of that would make it less ODD /
UNEXPECTED in its existence).
By all admission, ‘origin of life’
theories seem less plausible with each new, brilliant experiment and with each
further discovery of cellular processes. The optimism of the past—at being able
to synthesize life in the lab—has given way to a rather bleak pessimism (even
given the almost hubric optimism of good scientists-lol) of the main
researchers in the field.
We do not need to get into the
‘origin of life’ debate here—all we need to note (for my purposes here) is that
all the brilliant science research that has been done in this area has only
served to support the position that life (and most of the components inside the
cell) are “of inexplicable origin”… Consider just a few summary statements by
those working inside the field and in
adjacent fields.
Koonin makes a strong statement
about how difficult this issue is:
“In the preceding
chapter, we discussed possible scenarios for the origin of cells and
(hopefully) reached some degree of plausibility with the primordial Virus World
scenario of cellular evolution. However, this was all about relatively late stages
of evolution, at which replication of the genetic material and translation
yielding diverse proteins were already well established. All these models
seem to be of dubious value unless we develop some kind of explanation
for the origin of the fundamental processes of information transmission.
“The origin of life is the most difficult problem that faces
evolutionary biology and, arguably, biology in general. Indeed, the
problem is so hard and the current state of the art seems so frustrating
that some researchers prefer to dismiss the entire issue as being outside the
scientific domain altogether, on the grounds that unique events are
not conducive to scientific study. However, this position appears deeply
unsatisfactory, especially because, although life certainly evolved only once
on this planet (---), we have no idea just how unique (or otherwise) it is in
our universe as a whole. If one does accept the origin of life as a scientific
issue, then there seems to be no denying that it is a problem of overwhelming
importance before which all other questions in biology are relatively
mundane.” [p 351]
This is a fairly strong statement,
but the data seems to support it…
Results
not encouraging so far…
“Francis Crick once described the origin of life as ‘almost a miracle, so many are the
conditions which would have had to have been satisfied to get it going’.
And it’s true that the more ‘miraculous’ life appears to be, the harder it is to figure out how it can have started.
In 1859 Charles Darwin’s magnum opus On the Origin of Species first
appeared. In it he presented a marvellous account of how life has evolved over
billions of years from simple microbes to the richness and complexity of
Earth’s biosphere today. But he pointedly left out of his account the
question of how life got started in the first place. ‘One might as well
speculate about the origin of matter,’
he quipped in a letter to a friend. Today, we are not much further
forward.” [Davies, Demon, p. 167]
“Armed with a basic notion of the chemical setting (no
oxygen!), scientists have spent decades trying to re-create conditions
in the laboratory that might illuminate the first chemical steps on the long
pathway to life, following Miller’s pioneering efforts in 1953. Many subsequent
pre-biotic synthesis experiments have been done but, to be honest, they don’t
get very far, in spite of the dedication and ingenuity of the scientists. By the standards of biological molecular complexity,
these attempts barely make it to first base.”
[Davies, Demon,
p 172]
“Focusing on chemical correlates has yielded
limited progress in solving the origin of life, even after many decades of
effort.” [Walker, p. 50]
“RNA World hypothesis. This model proposes that life
started with RNA, as opposed to DNA (deoxyribonucleic acid), as the genetic
material. During the last few decades, the RNA World has been considered the
leading hypothesis for the origin of life on Earth. Gerry studies the in vitro evolution of RNA molecules to
understand potential mechanisms of chemical evolution in the putative RNA
World. He and his colleagues have demonstrated amazing capabilities in RNA,
such as the ability for self-replication and evolution under controlled
conditions. Because RNA chemistry isolated and evolved in a test tube can
fulfill so many functions of living biochemistry, it is understandable one
might arrive at a definition of life as a self-sustaining chemical system
capable of Darwinian evolution. A lot of people use this definition and like it.
But at the same time, like all definitions for life, it fails to deliver on
solving the hardest problems. It has not
allowed us to prove a mechanism for the origin of
life nor design new instruments
to search for and measure the presence of alien life on another world. … Let’s
first focus on the notion of “self-sustaining.” In vitro evolution
proceeds by taking a sample from one test tube and transferring it to
another, and then another, to simulate successive rounds of evolutionary
selection. Because evolution occurs only
by intervention of the experimenter, it requires continual human input to
sustain an evolving population of RNA. Most people do not feel
comfortable referring to these systems as “life” because they are not
self-sustaining and they require human intervention. Most people thus take
the stance, like the NASA working group did, that such systems should be
excluded.” [Walker, pp. 10-11]
“So, according to this view, it is easy to define
life: life is information that replicates itself. … If life is
information, where did this information come from initially? We asked this
question in chapter 7, and while it may appear at first that replacing the
question “What is the origin of life?” with “What is the origin of
information?” trivializes the search for our origins, it turns out that the
question is still hard. According to what we know today, the amount of information
necessary (and sufficient) to encode that information’s own
replication cannot simply emerge by chance. And assuming that the first
form of life arrived on Earth from
somewhere else in the solar system or galaxy does not solve the problem,
because the mathematics of the chance emergence of information does not change
if we move from one planet to another. This does not mean that the emergence
of life from nonlife is impossible, it just means that we have yet to
discover the pathway that allows information growth in a system where Darwinian evolution has yet to take
hold. …“ [Adami, p. 507]
“Selfishness cannot simply be assumed, however. That a chemical system developed the impetus
not merely to replicate, like a crystal, but to replicate specific
sequences of complex molecules and persist against existential threat, are phenomena that demand explanation. None of
the inorganic self-ordering, autocatalytic, dissipative structures known to
contemporary science does this (--). A crystal, a candle flame, a
hurricane, or a Benard cell does not seek
resources when the material conditions for continued catalysis
runs out; they cease. Living
things do so until all options are exhausted. Some of the simplest organisms
engage in surprisingly elaborate behaviors to forestall cessation.” [“To Be or Not To Be: Where Is
Self-Preservation in Evolutionary Theory?”.
Pamela Lyon. In Transitions, p. 105.]
“The uncritical assumption of selfishness creates a fundamental instability in much current
work on the evolutionary transitions and origins of life. Some of the
leading transitions narratives assume selfishness explicitly (Michod and Roze
1997) or implicitly (Maynard Smith and Szathmary 1995) at their foundation. All current
origin-of-life scenarios of which I am aware also appear to presume the
emergence of selfishness, or the kind of self-organized organized complexity
that supports the attribution of similar concepts, at a very early stage, well before
cells appear. Since this assumption is unsupported, origins
narratives remain incomplete until the emergence of self-preserving behavior is
explained.”
[“To Be or Not To Be:
Where Is Self-Preservation in Evolutionary Theory?”. Pamela Lyon. In Transitions,
p. 106.]
“The degree of empirical support for the RNA World
hypothesis is not central to my argument. However, Orgel’s
pessimistic estimation of the evidence supporting his hypothesis places
in useful perspective unarticulated assumptions about system-preserving
behavior in the development of life, so I will briefly outline it here. In
his last review of progress on the RNA World hypothesis before his death in
2007, Orgel reports that “there is at present no
convincing, prebiotic total synthesis of any of the nucleotides” necessary
to support RNA synthesis (Orgel 2004, 108; my italics). Further, no evidence
has yet emerged linking ribozymes, “the smoking gun that ... led to the more
general acceptance of the RNA World hypothesis,” to genetic transcription or
RNA replication (Orgel 2004, 113). “[A]biotic
synthesis of RNA is so difficult,” Orgel concludes, “that it is unclear that the RNA World could have evolved de
novo on the primitive Earth” at all (100), a conclusion first
advanced by Cairns-Smith 30 years ago.
… Thus, in the opinion of one of its leading
advocates, scientific investigation of
the RNA World hypothesis has yielded much interesting chemistry in the past 40
years but little progress toward an adequate
explanation for the origins of informational molecules, much less life. The
language Orgel uses to sketch “some consequences of the RNA World hypothesis”
is all the more notable, therefore, because an informational molecule is still
only a hope, not a reality. It is the sort of language used commonly in origins
and transitions narratives….”
[“To Be or Not To Be:
Where Is Self-Preservation in Evolutionary Theory?”. Pamela Lyon. In Transitions,
Page 110.]
“For all Darwin’s indisputable achievements, he left a sizable problem for future
generations to solve: namely, the question of how the first “primordial form, into which life was first breathed,”
from which “all the organic beings which have ever lived on this earth have
descended” – that “simple beginning” from which “endless forms most beautiful
and most wonderful have been, and are being, evolved” – first came into existence.”
[“Self-Organization,
Self-Assembly, and the Inherent Activity of Matter” in Transformations,
1st paragraph of the chapter.]
“However, this is about all the good news; the rest
is more like a sobering cold shower. For all the advances of
“ribozymology,” no ribozyme polymerase comes close
to what is required if we are to accept an RNA-only replicator system as
a key intermediate stage in the evolution of life. Nor are any ribozymes
capable of catalyzing the synthesis of nucleotides or even their sugar
moieties. Even sweeping all these problems under the proverbial rug, the path
from a putative RNA World to the translation system is incredibly steep. The
general idea of a function(s) for abiogenic amino acids and possibly peptides in
the RNA World, such as the role of ribozyme cofactors, appears fruitful and is
compatible with experimental data. Nevertheless, breaking the evolution of
the translation system into incremental steps, each associated with a
biologically plausible selective advantage, is extremely
difficult even within a speculative scheme let alone
experimentally. The triplicase / protoribosome hypothesis is attractive as an
attempt to explain the origin of translation and replication in one sweep, but
is this scenario realistic? The triplicase itself would have to be an extremely
complex, elaborate molecular machine, leaving one with the suspicion that,
all its attraction notwithstanding, the triplicase might not be the most likely
solution to the origin of translation problem. … All things considered, my assessment of
the current state of the art in the study of the origins of replication and
translation is rather somber. Notwithstanding relevant theoretical models and
suggestive experimental results, we currently do
not have a credible solution to these problems and do not even see with any clarity a path to such a
solution.” [Koonin, pp.
376-377].
“As pointed out earlier, the overall situation in
the origin of life field appears rather grim. Even under the (highly nontrivial) assumption
that monomers such as NTP are readily available, the problem of the
synthesis of sufficiently stable, structurally regular polymers (RNA) is formidable, and the origin of replication
and translation from such primordial RNA molecules could
be an even harder problem. … The emergence of the first replicator system,
which represented the “Darwinian breakthrough,” was inevitably preceded by a
succession of complex, difficult steps for which biological evolutionary
mechanisms were not accessible. Even considering environments that could
facilitate these processes, such as networks of inorganic compartments at
hydrothermal vents, multiplication of the probabilities for these steps could
make the emergence of the first replicators
staggeringly improbable.” [Koonin, p. 382]
”However, the origin of life—or, to be more
precise, the origin of the first replicator systems and the origin of
translation—remains a huge enigma, and progress
in solving these problems has been very modest—in the case of translation,
nearly negligible. Some potentially fruitful observations and ideas
exist, such as the discovery of plausible hatcheries for life, the networks of
inorganic compartments at hydrothermal vents, and the chemical versatility of
ribozymes that fuels the RNA World hypothesis. However, these advances remain
only preliminaries, even if important ones, because they do not even come
close to a coherent scenario for pre-biological evolution, from the first
organic molecules to the first replicator systems, and from these to bona
fide biological entities in which information storage and function are
partitioned between distinct classes of molecules (nucleic acids and proteins,
respectively). In my view, all advances notwithstanding, evolutionary
biology is and will remain woefully incomplete until there is at least a
plausible, even if not compelling, origin of life scenario.” [Koonin, p. 417]
“This information hierarchy—DNA to RNA to proteins—was
worked out in the 1950s-60s and allowed scientists to comprehend the general
mechanism by which the self-maintenance of organisms occurs. Nevertheless,
this understanding does not explain the origin
of such a mechanism or how such a system could evolve in the first place. By understanding the central dogma, we may
begin to imagine and test experimentally the potential prebiotic
chemistry that could have provided the foundation for the eventual emergence of
something so sophisticated. [Prebiotic, p. 41]
“mRNA molecules are produced using DNA templates via
transcription. mRNA is read out using ribosomes to make proteins via
translation. It remains unknown how
these processes originated.” [Prebiotic, p. 46]
Lab versus “messy earth”
“For the majority of the history of prebiotic
chemistry research (and organic synthesis as a whole), most reactions have been
carried out using stepwise protocols starting with pure reagents,
commensurate with the teachings and techniques for what might be termed
traditional organic chemistry. There is a growing recognition by the prebiotic
chemistry research community that geochemical
environments would have been ‘messy; leading to complex mixtures, and
that this complex chemistry, which may be crucial for the emergence of
life-like behaviours needs to be investigated experimentally.” [Prebiotic,
p. 6]
“The fundamental problem is not the simplicity of the
components in these experiments. It is something far deeper. To attain even
the modest successes announced so far requires special equipment and
technicians, purified and refined substances, high-fidelity control over
physical conditions — and a big budget. But above all, it needs an
intelligent designer (aka a clever scientist). The organic chemist must have a preconceived notion of the entity to be manufactured.
I’m not denigrating the scientists involved or the glittering promise of the
field of synthetic biology, only its relevance to the natural origin of life. Astrobiologists
want to know how life began without fancy equipment, purification procedures,
environment-stabilizing systems and — most of all — without an intelligent
designer. It may turn out that life is indeed easy to make in the lab but
would still be exceedingly unlikely to happen spontaneously in the grubby
and uncertain conditions available to Mother Nature. After all, organic
chemists can readily make plastics, but we don’t find them occurring naturally.
Even something as simple as a bow and arrow is straightforward for a child to
make, but would never be created by an inanimate process. So just because we
might (one day) find life easy to create does not of itself demonstrate a
cosmic imperative.” [Davies, Demon, pp. 178-179]
“There is a long history of what might be called
‘molecular Darwinism’ in which ‘naked’ molecules are able to replicate with
varying efficiency and natural selection filters out the best. The so-called RNA world theory falls into
this category. Though these studies are instructive, they are very
contrived and require carefully managed human intervention (e.g. to prepare
materials, to do the selecting) to accomplish anything. Relevance to the
natural world is far from obvious.” [Davies, Demon, p. 183, note 2]
“Ribonucleic acid (RNA) holds a special place in
origins-of-life research, because its sequence provides both a potential
mechanism for storing information as well as carrying out catalysis and other
functions. In extant biology, however, the synthesis of these biopolymers
including RNA requires a suite of sophisticated (highly evolved) proteins as
well as the ribosome, which a
priori would not have existed during the
early stages of prebiotic chemistry” [Prebiotic, p. 117]
The
Ratchet problem:
“As I have mentioned, a basic unknown
about the pathway from non-life to life is whether it was a long, steady slog
up a pre-biotic version of Mount Improbable, or whether it took place in fits
and starts, with long periods of stasis interrupted by great leaps forward (or
upward, in this metaphor). Given that Mount Improbable is so incredibly high, it
won’t do for a chemical mixture to attain a toehold in the foothills only to
slide back down again. There has to be some sort of ratcheting effect to
lock in the gains and limit the losses while the system hangs out for the
next step. But ideas like this, which seem sensible enough, run into the
problem of teleology. A chemical soup doesn’t know it’s trying to make life – a chemical soup doesn’t know anything at all – so it won’t act to protect its hard-won
complexity from the ravages of the second law of thermodynamics.
Scenarios in which chemistry ‘strives’ towards life are patently absurd. The same problem doesn’t
occur once life gets going, because natural selection can ratchet up the gains
and DNA storage can lock them in. But
chemistry without natural selection has no recourse to such mechanisms…” [Davies,
Demon, pp. 172-175]
“The backsliding problem afflicts almost
all studies of the complexification pathway to life. There are many clever
experiments and theoretical analyses demonstrating the spontaneous formation of
complexity in a chemical mixture, but they all hit the same issue: what
happens next? How does a chemical broth build on some spontaneously
emerging complexity to then ramp up
to something even more complex? And on and on, until the summit of the
pre-biotic Mount Improbable is reached? The most promising break-out from this
straitjacket comes from work on ‘autocatalytic’ chemical cycles. The idea here
is that certain molecules, say A and B, react to make other molecules, C, that
happen to serve as catalysts to accelerate the production of A and B. There is
thus a feedback loop: groups of molecules catalyse their own production. Scaling this up, there could be a vast
network of organic molecules forming a quasi-stable system of autocatalysis,
with many interlocking feedback loops, combining in a tangled web of reactions
that is self-sustaining and robust. All this is easy to state in words, but
are there such chemical systems out there? Yes, there are. They are called living
organisms and they deploy all the aforementioned features. But now we are
going round in circles, because we want to ascertain how all this marvellous
chemistry can take place before life. We can’t put the solution by hand into
the problem we are trying to solve and then claim to have solved it.” [Davies, Demon,
pp. 172-175]
“Multiple rounds of prebiotic RNA
template-directed synthesis still remain a
challenge but are necessary in order to have a system capable of
sustained replication and hence Darwinian evolution. The biggest obstacle
to achieving this is the fact that a successful round of template copying is
prone to result in a stable double-stranded structure. In order for another
round of template-directed synthesis to begin, the two strands must be
separated and remain separated long enough for on-template polymerization/ligation to take place. The
fact that the resulting duplex impedes further
rounds of copying is known as the strand inhibition problem.
Two hybridized strands can potentially be separated simply by raising the temperature
past the melting points; however, high temperatures also prevent Watson-Crick
pairing of the activated mono- and oligonucleotides.” [Prebiotic, p. 132]
“Another rather ingenious strategy for
achieving multiple rounds of RNA synthesis takes advantage of chimeric
templates… however, the resulting RNA-chimeric duplex is not as stable
as the corresponding pure RNA duplex. This weaker duplex means the chimeric
strand can be readily displaced by two, smaller complementary RNA
oligonucleotides…. Achieving multiple rounds of RNA copying still has
challenges to be overcome but is currently an active area of research.
Success would open the door to producing chemical systems which can begin
to undergo Darwinian evolution in the laboratory in a potentially prebiotic
manner.” [Prebiotic, p. 132]
“The Fitness
Landscape before Evolution The central
problem that we have to surmount when it comes to understanding the origin of
life is that there appears to be an
insurmountable gap between the minimum
amount of information it takes to self-replicate, and the amount of information that can appear by chance,
even if we account for a biased monomer
distribution that can deliver a significant amount of information “for free,” as it were, via the enhancement
factor the bias provides. In order to bridge this gap, we need to uncover a
dynamical process that will drive
information to higher levels in a non-equilibrium fashion, perhaps
in a ratchet manner (---). To this
day, no one has identified such a process. But even if we could identify
it, can such a process reach the flanks of
the fitness peaks of the primordial fitness landscape: the landscape
of the simplest self-replicators?” [Adami, p. 403]
Chicken or Egg problem
“Obviously, when
considering the origin of the first life forms, one faces the proverbial
chicken-and-egg problem: What came first, DNA or protein, the gene or the
product? In that form, the problem might be outright unsolvable due to
the Darwin-Eigen paradox: To replicate and transcribe DNA, functionally active
proteins are required, but production of these proteins requires accurate
replication, transcription, and translation of nucleic acids. If one sticks
to the triad of the Central Dogma, it is impossible to envisage what
could be the starting material for the Darwin-Eigen cycle. Even removing DNA
from the triad and postulating that the original genetic material consisted of
RNA (thus reducing the triad to a dyad), although an important idea (---), does
not help much because the paradox remains. For the evolution toward greater
complexity to take off, the system needs to somehow get started on the
Darwin-Eigen cycle before establishing the feedback between the (RNA) templates
(the information component of the replicator system) and proteins (the
executive component).
[Koonin, p. 360]
“The crucial
question in the study of the origin of life is how the Darwin-Eigen cycle started—how was the minimum complexity that is required
to achieve the minimally acceptable replication fidelity attained?
In even the simplest modern systems, such as RNA viruses with the replication
fidelity of only about 10(-3) and viroids that replicate with the
lowest fidelity among the known replicons (about 10(-2) ---),
replication is catalyzed by complex protein polymerases. The replicase itself
is produced by translation of the respective mRNA(s), which is mediated by
the immensely complex ribosomal apparatus. Hence, the dramatic
paradox of the origin of life is that, to attain the minimum complexity
required for a biological system to start on the Darwin-Eigen spiral, a system of a far greater complexity appears to be
required. How such a system could evolve is a puzzle that
defeats conventional evolutionary thinking, all of which is about biological
systems moving along the spiral; the solution is bound to be unusual.” [Koonin,
p. 353]
“In living things,
most chemical activity is handled by proteins. Metabolism — the flow of energy
and material through organisms — is necessary for life to achieve anything, and
proteins do the lion’s share of metabolic work. If life got started (as
Prigogine believed) via elaborate energy-driven chemical cycles, then proteins
must have been early actors in the great drama of life. But on their own,
proteins are largely useless. The all-important organization of life
requires a great deal of choreography, that is, some form of
command-and-control arrangement. That job is done by nucleic acids (DNA and
RNA). Life as we know it involves a deal struck between these two very
different classes of molecules: nucleic acids and proteins. The conundrum
as most scientists see it is the chicken-and-egg
nature of life: you can’t have one without the other. Without a
legion of proteins to fuss around it, a molecule of DNA is stranded. In simplistic
terms, the job descriptions are: nucleic acids store the details about the ‘life
plan’; proteins do the grunt work to run the organism. Both are needed.”
[Davies, Demon, p. 22]
”The “chicken—egg
relationship” between genes and proteins: The so-called “chicken—egg
relationship” has made it difficult to solve the mystery of the origin of life
for many years. In such a situation, the RNA world hypothesis, which assumes
that first life arose from the RNA world, which was formed by the
self-replication of RNA, was proposed by Gilbert in 1986 [---]. However, it
would be principally impossible to solve the from the standpoint of this
hypothesis because any gene encoding a mature protein could never be formed in
the absence of a target protein or in the absence of protein, even if RNA could
be produced on primitive Earth.” [Ikehara, K. “How Did Life Emerge in
Chemically Complex Messy Environments?” Life 2022, 12, 1319]
“So an inevitable (even if perhaps
counterintuitive) conclusion from the comparative analysis of ancient
paralogous relationship between protein components of the translation system is
that, with the interesting exception of the core ribosomal proteins, all
proteins that play essential roles in modern translation are products of a long
and complex evolution of diverse protein domains. Here comes the Catch-22: For all this protein evolution to occur, an accurate and
efficient translation system is required. This primordial
translation system might not need to be quite as good as the modern version,
but it seems a safe bet that it must have been within an order of magnitude
from the modern one in terms of fidelity and translation rates to make protein
evolution possible. However, from all we know about the modern translation
system, this level of precision is unimaginable without a complex, dedicated
protein apparatus…. Thus, the translation system presents us with the “Darwin-Eigen
paradox” that is inherent to all thinking on the emergence of complex
biological entities: For a modern-type, efficient, and accurate translation
system to function, many diverse proteins are
required, but for those proteins to evolve, a translation system almost as good
as the modern one would be necessary. There
seems to be only one conceivable solution to this paradox—namely, a (partial)
refutation of the first part of the opposition: We are forced to conclude that
a translation system comparable to the modern one in terms of accuracy and
speed functioned without many proteins, possibly without any proteins at
all. Hence, the existence of a complex, elaborate RNA World, in which a
primitive version of the Darwin-Eigen cycle was already operating, can be
conjectured from the comparative analysis of the translation system components.”
[Koonin, p. 359]
(A special version
of the chicken-and-egg problem occurs with eukaryotes. Certain error-control
systems had to be in place when euk’s appeared (“co-evolved”), but do not show
up in other life forms:
“As we discussed
in great detail in Chapter 7, eukaryotes possess an important
information-processing step that effectively has no counterpart in
prokaryotes: splicing of primary transcripts. The accompanying
quality control system apparently has evolved concomitantly with
eukaryogenesis: the nonsense-mediated decay (NMD) machinery which
recognizes and destroys aberrant mRNAs that contain in-frame stop codons in
exons other than the last, 3’-terminal exon of the coding sequence (---).” [Koonin,
pp. 279f]
And there were ‘hostile’
elements working AGAINST even getting started…
Even the elements
important to life’s existence noted in the earlier installment, can WORK
AGAINST any putative pre-biotic processes thought to be involved in life’s
emergence.
For example, oxygen
– critical to much of life’s metabolism on earth today – destroys most things.
And a ‘reducing’ atmosphere – believed by many OOL (Origin Of Life) researchers
to be a requirement for the origination of primitive life– would kill most
air-breathers today. So, Davies [Demon, pp. 170-171]:
“Shifting the
cradle of life from Earth to somewhere else doesn’t much advance the more
important question of what geological setting would be conducive to producing
life. Many scenarios have been touted: deep ocean volcanic vents, drying
lagoons, pores in sub-ocean rocks ... the list is long. About the only thing
everyone agrees on is that oxygen gas would have been a frustrating factor.
Today, complex organisms require oxygen for their metabolism, but this was
a late development. On Earth, there was very little free oxygen in the
atmosphere before about 2 billion years ago, and present levels were not
attained until within the last billion years. Oxygen may feel good to
breathe, but it is a highly reactive substance that attacks and breaks down
organic molecules. Aerobic life has evolved all sorts of mechanisms to cope
with it (such as anti- oxidants). Even so, reactive oxygenic molecules
regularly damage DNA and cause cancer. When it comes to the origin of life,
free oxygen is a menace. “
And water – so
essential to life as we know it – would have been a major ‘frustrating factor’
in pre-biotic processes, since it tends to ‘dissolve stuff.
“Life as we know
it would not exist without water. However, water molecules not only serve as a
solvent and reactant but can also promote hydrolysis, which counteracts the formation of essential organic
molecules. This conundrum constitutes one of the central issues in origin
of life. Hydrolysis is an important part of energy metabolism for all living
organisms but only because, inside cells, it is a controlled reaction.” [“The
ambivalent role of water at the origins of life”, Vieira, Kleinermanns, Martin,
Preiner. FEBS Lett. 2020 Sep]
“The “water
paradox” is a crucial problem in the research on the prebiotic chemical
evolution towards the emergence of life: It states that although aqueous
environments are essential for life, they impede key chemical reactions such
as nucleotide polymerization. In
aiming to overcome this paradox various hypotheses have been proposed,
including scenarios based on alternative solvents like formamide, condensing
agents like cyanamide, high temperatures (~160°C) or approaches based on
wet/dry cycles. However, when appraising the prebiotic plausibility of such
scenarios general weaknesses appear. Besides the fact that all known life
manages the water paradox without needing such proposed conditions,
evolutionary conservatism – the principle that evolution builds on existing
pathways – indicates that the same physicochemical effects were probably
involved in the abiotic origin of biopolymers as now being tapped by life via
complex enzymes.” [“Natural Nanofluidic Environments as Prebiotic Reaction
Vessels: Abiotic RNA formation in temporal nanoconfined water”, Frank Trixler, NORCEL
Institute.]
But not only does
water ‘IMPEDE” formation, but it REVERSES it too:
“The fact that
water favours the decomposition of RNA and
peptides back into their monomer components, as well as other
hydrolysis reactions in prebiotic chemistry, is sometimes referred to as the
water problem. There is every indication, however, that water is necessary for
life, but at the same time, water is constantly working against polymerization.”
[Prebiotic, pp. 3-4]
A special case is
the ‘space chemistry, where it has been argued that bio-active molecules
arrived from space—to jump start life:
“Finally, the
results obtained from the examination of the space
chemistry should be summarized. The most conspicuous negative
result is the fact that numerous compounds
that play a fundamental role for a successful CE were not found.
Here, the following compounds should be mentioned: several prAAs such as Arg,
His, and Cys, and oligopeptides; furthermore, the nucleobase cytosine, any kind
of nucleosides, nucleotides, and oligonucleotides; last but not least, unsubstituted
long alkanols or alkanoic acids which are needed for the formation of lipid
membranes are extremely rare compared to the methyl substituted analogs. Even,
when traces of some of these compounds will be detected in the future, such
findings will not change the conclusion that the space chemistry was and is far from supporting a CE. This conclusion
is confirmed by the fact that the space
chemistry creates large amounts of chemicals that are unfavorable or even a
definite obstacle for a CE. Such compounds are large amounts of
non-prAAs and imino acids, large amounts of formic acid, alkanoic acids,
hydroxy acids, and even sulfonic acids.”
[Kricheldorf, Hans
R.. Life by Chemical Evolution?: A Review and Evaluation of Experiments and
Hypotheses (pp. 417-418). Springer Berlin Heidelberg. Kindle Edition.]
But these are
attacks on ‘life’ before it gets started, and there are many, many more
attacks ONCE ITS HERE (e.g. multiple mass extinctions of early life on our
planet) to talk about later.
…………………………………..
But even if this trend reversed
eventually, and they could do it in a seriously constrained lab with zero
contaminants, and incredibly precise control operations, that would be (1)
unrepresentative of earth conditions as far from “lab-like” conditions as
possible ; (2) require all these test tubes to be co-located and
simultaneously/sequentially run, and (3) STILL not reduce the shock from
the ‘how is it STILL HERE’ (given all the assaults on it throughout early earth
history) question.
The most recent (and brutally
honest) assessment/review of all the chemical experiments and hypotheses that I
have found is Life by Chemical Evolution?: A
Review and Evaluation of Experiments and Hypotheses, Springer:2025. By Hans R. Kricheldorf. (of the
Institute for Technical and Macromolecular Chemistry, Hamburg Germany).
After giving the history of the
demise of the ‘prebiotic soup’ hypothesis, he makes this observation about its
‘resistance to decay’, in his summary chapter:
“Nonetheless, it
is amazing that even after the year 2000, the prebiotic soup hypothesis still
plays a role as standard scenario for a CE, which yielded a living cell
starting out from a mixture of simple molecules such as CO2, N2,
H2, and H20. This view is above all characteristic for
biologists as may be demonstrated by a comment of Ribas de Pouplana in his book
“The genetic code and the origin of life” (Ribas de Poublana 2004):
“The most widely
accepted scenario for the transition from abiotic to biotic chemistry is that
the simple monomeric compounds present in the prebiotic soup somehow underwent
polymerization, perhaps with the assistance of clays and minerals and formed
longer and longer chains or polymers, which over time became increasingly more
complex with respect to both their structure and their properties.”
In this way, the
wonder of the primordial soup as the birthplace of life will be preserved for
eternity, at least for laymen of chemistry.
He points out later that there is
no reason to believe that CE (Chemical Evolution) happened:
“Finally, the
fundamental problem should be discussed, namely the question, why a CE on the
early Earth has resulted in the formation of a living cell. Even, when the
number of model reactions demonstrating how individual steps of the CE might
have occurred will further increase, the knowledge
of hundreds of individual reactions does not necessarily explain, why a CE
followed the road map toward a living cell. Increasing complexity of
groups of molecules also means increasing diversity for the entire pool of
organic compounds on Earth, but the driving force for an increasing diversity
is a gain of entropy. This correlation is best and clearly demonstrated by the “space
chemistry.” However, a living cell practices a different strategy, it maintains
thermodynamically instable, kinetically controlled processes and compounds
against a gain of entropy by permanent consumption of (sun) energy.”
He makes the point that belief in
the confluence of all factors at the same time
(against the evidence) is essentially a ‘hypothesis of a fortunate
coincidence’—and not a scientific theory—especially when contraindicated by
polymer chemistry:
“The hypothesis of
a fortunate coincidence or contingency is for three reasons not an acceptable
hypothesis of CE.
“First, scientific
research concerns discovery evaluation and application of laws of nature. A law
of nature is evident from the reproducibility of (experimental) observations
which are reproducible independently of time, location, and researcher. Yet, singular
events are per definition not part of a law of nature, and thus outside the
reach of scientific methods.
“Second, it has
not been demonstrated yet that all the
components needed for the formation of a living were simultaneously present on
the early Earth, regardless whether the primordial soup or a
compartment concept is taken into account. The
experiments and observation listed in the preceding chapters rather suggest
that a more or less simultaneous evolution of homochiral proteins, RNAs, and
amphiphilic molecules forming stable vesicles was not possible.
When the RNA world hypothesis is taken into account and when an independent
evolution of proteins is ignored, one still needs a strong belief in
chemical miracles to assume that those RNAs needed for the production of
bioactive proteins were formed in significant quantities, came together and
recognized that they had to cooperate for the production of useful proteins.
“Third, it
requires again a strong belief in wonders to expect that the problem of
homochirality was solved by a chance event. Even when one concedes that a
fortunate event has accumulated homochiral biopolymers, the racemization
would have destroyed the homochirality within a short time except at
temperature around or below 0 degrees C.”
When he get gets around to stating
his position, he is blunt about what he sees the data pointing to:
“The reader would
presumably like to know what is final conclusion of the author. In short, the
author is not able to reach a final decision. The hypothesis of a CE is a
fascinating vision which (as pointed out below) allows for a consistent
interpretation of the structure of the universe, but there are so many open
questions and unsolved fundamental problems that the
author is not convinced yet that a CE beginning with simple gases and
ending up with a living cell has really taken place.”
He points out that researcher
optimism about CE is more ‘religious’ or ‘ideological’ than scientific:
“A researcher, who
is a priori convinced that the hypothesis of CE is true, he takes an ideological
stance and not a scientific one. Undirected evolution in combination with
chance events is not a scientific approach
to the explanation of a CE (because reproducibility is fundamentally
impossible) and requires so much belief in
miracles as any religion. The hypothesis of a directed evolution is
confronted with the question who was the “director,”, or which property of the
universe has taught the molecules to follow the pathway to a living cell. This
problem is acerbated by the finding that the reductionistic-deterministic
concept is not correct as evidenced by the failure of
numerous model experiments and by the course of the “space chemistry.”
And he states that any
non-religious approach has to accept (and explain?) the occurrence of TWO “Big
Bangs”—
“When the
existence of a CE is denied, again a religious and a non-religious answer is
possible. The religious answer is the creation of the first cell by a God
(Allah, Manitou, etc.). The non-religious view is to say that the design of
the universe includes two “Big Bangs,” the first one resulting in the
appearance of space, time energy and matter, and a second
“Big bang” resulting in the appearance of life with the first cell.”
__________________________________________________________
__________
FOUR:
… Nor how life is STILL here!
Before the modern era, life has
been assaulted multiple times, with devastating results.
What are called the ‘mass
extinctions’ eliminated species and large segments of all life.
“Among the most
striking features of the fossil record are the periods of accelerated mass
extinctions followed by periods of accelerated mass “originations” (appearances
of morphologically novel organisms).
The most famous of these episodes is the
event that led to the disappearance of dinosaurs at the end of the Cretaceous
period 65 MYA. … These mass extinctions occurred at the boundaries between one
geological age with its characteristic fauna and another with a different
fauna. Raup and Sepkowski first documented the “big five” mass extinctions in
1982:
“Since then, mass
extinctions have been identified at other major geological boundaries:”
[Shapiro, pp. 407-408]
The amount of damage estimated from
the largest of these is staggering:
One website listed 44 extinction
events (of various scales—not all of which are comparable):
https://www.nazaudy.com/list-of-extinction-events
Extinction 1 –
(3.8 Gya) Archean Eon and the Carbon Genesis
Extinction 2 –
(2.4 Gya) Huronian Glaciation, Snowball Earth 1 – Great Oxygenation Event (GOE)
– First major Ice Age
Extinction 3 –
(650 Mya) Stuartian Glaciation, Snowball Earth 2 – Second Ice Age
Extinction 4 –
(542 Mya) End-Ediacaran Extinction
Extinction 5 –
(513-517 Mya) End-Botomian Extinction
Extinction 6 –
(502-497 Mya) Dresbachian Extinction
Extinction 7 –
(488 Mya) Cambrian-Ordovician Extinction
Extinction 8 –
(450 Mya) 86% of species lost: (1) Ordovician-Silurian Extinction
Extinction 9 –
(428 Mya) Ireviken event
Extinction 10 –
(424 Mya) Mulde event
Extinction 11 –
(420 Mya) Lau event
Extinction 12 –
(375 Mya) 86% of species lost: (2) Late Devonian Extinction episode
Extinction 13 –
(305 Mya) Carboniferous Rainforest Collapse
Extinction 14 –
(299 Mya) The Lignin breakthrough and demise
Extinction 15 –
(272 Mya) Olson’s Extinction
Extinction 16 –
(260 Mya) End-Capitanian extinction event
Extinction 17 –
(250 Mya) 96% of species lost: (3) The Great Permian Extinction
Extinction 18 –
(232 Mya) The Carnian Pluvial Episode (CPE)
Extinction 19 –
(200 Mya) 80% of species lost: (4) End of Triassic Extinction
Extinction 20 –
(183 Mya) Toarcian turnover
Extinction 21 –
(145 Mya) End-Jurassic (Tithonian) extinction
Extinction 22 –
(117 Mya) Aptian Extinction
Extinction 23 –
(94 Mya) Cenomanian-Turonian boundary event
Extinction 24 –
(66 Mya) 76% of species lost: (5) End of Cretaceous and the Dinosaurs
Extinction 25 –
(56 Mya) Paleocene-Eocene Thermal Maximum (PETM)
Extinction 26 –
(33.9 Mya) Eocene–Oligocene Transition event (EOT)
Extinction 27 –
(16.9 Mya) Mid Miocene Climate Optimum (MMCO)
Extinction 28 –
(14.5 Mya) Middle Miocene disruption
Extinction 29 –
(5.96 Mya) Kapitean-Optian Glacial Event
Extinction 30 –
(5.3 Mya) The Zanclean flood
Extinction 31 – (3
Mya) The Great American Biotic Interchange
Extinction 32 –
(2.6 Mya) Pliocene–Pleistocene boundary extinction
Extinction 33 –
(2.1 Mya) Yellowstone eruption number 1
Extinction 34 –
(1.3 Mya) Yellowstone eruption number 2
Extinction 35 –
(1.09 Mya) The Gakkel Ridge Caldera
Extinction 36 –
(790,000) The Bolaven meteor impact
Extinction 37 –
(640,000) Yellowstone eruption number 3
Extinction 38 –
(400,000) The Homo species extinctions and fire
Extinction 39 –
(130,000 years ago) The Eemian interglacial
Extinction 40 –
(74,000 years go) The Toba eruption
Extinction 41 –
(46,000 years ago) Quaternary extinction, Australian Megafauna disappearance
Extinction 42 –
(39,000 years ago) Campanian Ignimbrite eruption
Extinction 43 – (26,500
years ago) Oruanui eruption
Extinction 44 – (24,000
years ago) end of Neanderthals
Extinction 45 –
(14,000 years ago) Quaternary extinction, North America Megafauna disappearance
Extinction 46 –
(13,000 years ago) Lake Missoula drainage
Extinction 47 –
(12,000 years ago) The Younger Dryas
Extinction 48 –
(11,500 years ago) The Sundaland Submerge
Extinction 49 –
(11,000 years ago) Quaternary extinction, South America Megafauna disappearance
Extinction 50 –
(10,000 years ago) Quaternary extinction, Eurasia Megafauna disappearance
Extinction 51 –
(8,200 years ago) The sinking of Doggerland
Extinction 52 – (8,200
years ago) The collapse of Lake Agassiz
Extinction 53 –
(8,000 years ago) The flooding of the Persian Gulf basin
Extinction 54 –
(7,600 years ago) The Black Sea deluge
Extinction 55 – (4,000
years ago) The dry of the Saharam Forest
And yet ‘life’ somehow continued
and even became more robust and more diverse:
“Originations of new groups and rapid diversification
(or “radiation”) of certain groups, like Bilateria in the Cambrian, characteristically follow mass extinctions [--].
These rapidly radiating groups may have arisen in the previous geological
period, as we now know was the case for Bilateria [--]. There is general
agreement that depopulation of existing ecological niches and the creation of
new ecological niches by the novel organisms themselves are critical but hard
to quantify factors in the dynamics of the appearance of new organisms in the record.
Over time, of course, originations have to outnumber extinctions for life to
sustain and increase its necessary diversity. There is a long-term increase
in the number of taxonomic families visible in the fossil record that extends
over multiple mass extinction-origination episodes [--].” [Shapiro (pp.
408-409).]
But
the adaptability of life forms – wherever it comes from – has overcome so many
adverse events… and the ability of ‘primitive cells’ to survive and thrive in
extreme environments and using ’unusual’ energy sources (e.g. Chernobyl
radiation!) makes them so much better than us (lol)…
…………………………….
…………………..
There
are more ‘gaps’ in this area of life/biology (some of which are substantial—see
quote below), but they only ADD to the ‘level of shock’ of there being a
‘biosphere from nothing’.
So,
for example, Koonin [pp. 171ff]–
“Organisms with large, complex
cells are known as eukaryotes—that is, possessing bona fide
nuclei. These organisms include the three kingdoms of multicellular life forms,
plants, brown algae, and animals, as well as a huge variety of
unicellular forms (also known as protists).
Eukaryotic cells typically are orders of magnitude bigger than prokaryotic
cells and possess complex intracellular organization with diverse
membrane-bounded organelles, including the eponymous nucleus and the
mitochondria that evolved from endosymbiotic bacteria. Thus, by any reasonable criteria, eukaryotic cells are
dramatically more complex than bacteria and archaea. How this complexity evolved is a major enigma
of evolutionary biology, ……………………………………………
“It is neither practical nor
necessary in this book to delve into the innumerable fine details of biological
structures. However, to discuss the origin of eukaryotes (hereafter eukaryogenesis)
in earnest, we need to fully appreciate the
nature and depth of the gulf that separates the cells of eukaryotes from
prokaryotic cells. Indeed, there
is a sharp divide in the organizational complexity of the cell
between eukaryotes and prokaryotes: A typical eukaryotic
cell is about a thousand-fold larger by volume than a typical
bacterium or archaeon and possesses elaborate
intracellular compartmentalization that is not seen even in the most
sophisticated prokaryotes.
“The compartmentalization of
eukaryotic cells relies on an elaborate, diversified endomembrane system and
the actin-tubulin-based cytoskeleton. A striking consequence of the
intracellular compartmentalization is that eukaryotic cells are physically
distinct from the cells of prokaryotes. In prokaryotes, the content of the
cell is a solution, even if a viscous one, so macromolecules (proteins and
nucleic acids) diffuse more or less freely and reach their cellular
destinations through the combination of stochastic movements with trapping in
specific complexes. By contrast, in eukaryotes, macromolecules are
largely prevented from free diffusion and instead reach their target sites
through complex trafficking systems. … Proteins and nucleic acids ooze out
of permeabilized bacterial cells, but not (typically) from eukaryotic cells
(Hudder, et al., 2003). So the cytosol of eukaryotic cells has much lower
entropy than prokaryotic cells—it is hard to
think of a more fundamental difference.
“ Although the picture of
prokaryotic expression regulation is becoming increasingly complex and is
now a far cry from the simple Jacob-Monod scheme (see Chapter 5), nothing in prokaryotes can compare with the complexity of
the eukaryotic chromatin.
“Qualitative distinctions
between eukaryotes and prokaryotes are numerous and span various
aspects of cell biology, particularly those that are involved with
information processing, signal transduction, and intracellular trafficking
(see Box 7-1). The organizational complexity of the eukaryotic cells is
complemented by extremely sophisticated, cross-talking signaling networks.
“No direct counterparts to the signature eukaryotic
organelles, genomic features, and functional systems exist in archaea or
bacteria.
Hence, the very nature of the evolutionary relationships between prokaryotes
and eukaryotes becomes a cause of bewilderment. Indeed, sequence
comparisons between genome-wide gene sets show beyond doubt that a few thousand
eukaryotic genes responsible for key cellular functions (translation,
transcription, and replication) share common origins with homologs from archaea
and/or bacteria (Koonin, et al., 2004). This evolutionary unity of cellular
life forms makes it an extremely hard
and fascinating challenge to explain how largely common components give rise
to cells that are so dissimilar in so many respects.
But
we have to stop here (and do job-hunting), and start on the next
shock-factor: the social and community lives of these tiny organisms,
and their interactions with OTHER tiny organisms…
…………………………
The
mystery of life seems different than earlier ‘scientific mysteries’. Often the
scientist admits that something is ‘poorly understood’ (almost every article
abstract contains such a phrase, but the article tries to address it…) but
still states the mantra of science’s eventual conquest.
I
consider Shapiro to be a truly honest man, and even in the midst of
‘confession’ he can hold the line on the juggernaut of science:
“In order to be truthful, we must
acknowledge that certain questions, like the origins of the first living cells,
currently have no credible scientific answers. However, given the historical
record of science and technology in achieving the impossible (e.g., space
flight, telecommunications, electronic computation, and robotics), there is
no reason to believe that unsolved problems will remain without naturalistic
explanations indefinitely.” [Shapiro, p. 93]
But
I am convinced that this area will confound that… as some have already
admitted, after decades and decades of work:
“As Erica Hayden confessed in the journal Nature,
“As sequencing and other new technologies spew forth data,” the complexity
unearthed by cell biology “has seemed to grow by orders of magnitude.
Delving into it has been like zooming into a Mandelbrot set… that
reveals ever more intricate patterns as one peers closer at its boundary.” [Denton,
Miracle, p. 16]
“The origin of
life is the most difficult problem that faces evolutionary biology and, arguably,
biology in general. Indeed, the problem is so hard and the current state of
the art seems so frustrating that some
researchers prefer to dismiss the entire issue as being outside the scientific
domain altogether, on the grounds that unique events are not
conducive to scientific study.” [Koonin, p. 351]
[The
quantum substrate of reality may also be in this category, since it is
perfectly clear in the mathematics, but generally unclear in everything else..
almost koan-like (“Is the cat dead or alive? No, it quantum-tunneled into some cardboard
box.”…lol). And—just to compound ‘unfathomableness’ with more
‘unfathomableness’-- biological organisms USE quantum effects ALLTHE TIME …(e.g.
quantum tunneling in enzyme catalysis, olfactory and visual sensing,
photosynthesis, magnetoreception, DNA change—the whole field of Quantum
Biology…]
==================
It should be starkly obvious by now
that this ‘ball of teeming, churning life is odd
in the extreme—when compared to the ‘rest of the SOMETHING’—and even
more UNEXPECTED as appearing abruptly and without an
explanation…
In other words, it makes NO SENSE why it is here, and why it
is SO DEMONSTRABLY DIFFERENT from anything else in our experience of the
physical cosmos.
It is enigmatic in every dimension
of its existence and persistence.
Its ceaseless churning, and being
‘busy doing stuff’, and ‘acting like it knows what it is doing’ could not
have been foreseen, if we were just looking at the miniscule pile of chemicals,
or at life-less rocks and oceans…
Life sits either [on, under,
inside, around, over, enwrapping—or all of the
above] a batch of chemicals, and we would NOT have been able to ‘predict’ what
that batch of chemicals might become—once it was alive… There are no
trendlines, intermediate forms, analogous reactions to suggest that life would
show up and that it would be so incredibly sophisticated and competent in its
actions and choices.
This seems to be another case of
‘something from nothing’ (i.e. unknown causes?)--
Like the previous points, it is
almost scary to realize that there is NO NATURAL or SCIENTIFIC explanation for
its existence, exuberance, or expansiveness.
It is just ‘in our face’,
challenging us with the obvious intuition/implication of something ‘Outside’
that we might have to face someday in our conscious future…
The scary-factor comes from what it
might suggest about a putative OUTSIDE-Other…
We have already
conjectured/suspected that this Outside-Other was an expert mathematician,
physicist, cosmologist, logician, and chemist – to me as an
emotionally-stunted, socially-immature, OCD-ish, and artistically-monochromed –
this is not really threatening from a personality perspective.
The power/energy/competence
required to CREATE such a SOMETHING is – of course – VERY THREATENING.
But this new layer – that of life
that acts “like it knows what it is doing” (with types/levels/aspects of
cognition at all levels of hierarchy it participates in) and that forces
interaction with it—is NOT in my ‘comfort zone’. These things have
purpose/goals, and as we move up the taxonomic web, we find things (agents)
that look a lot like ME (e.g., domestic pets, service animals) in behavior and
aging and even learned habits. They provoke emotions (affections, fear), inspire
questions about ethics, and develop loyalty that transcends a reciprocity-only
relationship.
I know full well of the rapid life-death
cycle in them (with predation etc.), but overall, I consider life-as-a-whole to
be HEALTHIER than I am…
Where this hits me (in this
context) can be illustrated by my experiences in darker times in high school. I wrote literary things for the
school newspaper (and won an award for some of them – lol), but I could easily
recognize the robustness, health, vivacity of things written by ‘healthier’
people – people I actually ‘feared’ psychologically (for no reasons that
came from them—they were always nice to me). They were whole, balanced,
comfortable with emotions and uncertainty—I was not… I was defective,
less-than-alive, mutated, stunted… without doing ANYTHING, they were a constant
reminder (and a painful one) of my desiccated existence… [Many of you readers
will have had the same experience, I know…sigh]
And when I look NOW at this
exuberant and beautiful life-sphere under discussion [like their beautify and
wholesome literary works back then], I can sense the ‘wholeness and vivacity’
of whatever sourced it… this OUTSIDE-OTHER now stands apart from me—not
only in power, competence, and intelligence (of some kind)—but also in those
areas I am NOT such.
And so it would be naïve in
the extreme for me to presume that any post-mortem encounter with this
life-affirming/supporting putative Outside-Other would invariably be a ‘pleasant
experience’ for a disembodied mind (in the post-mortem state). Even if we could
confidently infer a ‘love of life’ to an OUTSIDE-OTHER, that might NOT be a
good/comforting thing, since such an Outside-Other might hold us accountable
for how we TREATED life-forms in our days upon the earth (e.g. some type of
judgment or consequences?).
Without some self-disclosure, all
our estimates of this Outside-other remains speculative—and we have no a-priori reason to believe in something
beneficent* (not all expert mathematicians, physicists, and
chemists--and universally and continuously ‘beneficent’).
[*Strictly speaking, my statement
here can be challenged philosophically—on a couple of points, coming up mostly
later in the exploration—but those do not carry the evidential force for what
we have discussed so far.]